Solanaceae
| Solanaceae | |
|---|---|
 | |
| A flowering Brugmansia suaveolens from the US Botanic Garden | |
| Scientific classification | |
| Kingdom: | Plantae |
| (unranked): | Angiosperms |
| (unranked): | Eudicots |
| (unranked): | Asterids |
| Order: | Solanales |
| Family: | Solanaceae Juss. |
| Subfamilies | |
|
Cestroideae | |

The Solanaceae, or nightshades, are an economically important family of flowering plants. The family ranges from annual and perennial herbs to vines, lianas, epiphytes, shrubs, and trees, and includes a number of important agricultural crops, medicinal plants, spices, weeds, and ornamentals. Many members of the family contain potent alkaloids, and some are highly toxic, but many cultures eat nightshades, in some cases as staple foods. The family belongs to the order Solanales, in the asterid group and class Magnoliopsida (dicotyledons).[2] The Solanaceae consists of about 98 genera and some 2,700 species,[3] with a great diversity of habitats, morphology and ecology.
The name Solanaceae derives from the genus Solanum, "the nightshade plant". The etymology of the Latin word is unclear. The name may come from a perceived resemblance of certain solanaceous flowers to the sun and its rays. At least one species of Solanum is known as the "sunberry". Alternatively, the name could originate from the Latin verb solari, meaning "to soothe", presumably referring to the soothing pharmacological properties of some of the psychoactive species of the family.
The family has a worldwide distribution, being present on all continents except Antarctica. The greatest diversity in species is found in South America and Central America.
The Solanaceae include a number of commonly collected or cultivated species. The most economically important genus of the family is Solanum, which contains the potato (S. tuberosum, in fact, another common name of the family is the "potato family"), the tomato (S. lycopersicum), and the eggplant or aubergine (S. melongena). Another important genus, Capsicum, produces both chili peppers and bell peppers.
The genus Physalis produces the so-called groundcherries, as well as the tomatillo (Physalis philadelphica), the Cape gooseberry and the Chinese lantern. The genus Lycium contains the boxthorns and the wolfberry Lycium barbarum. Nicotiana contains, among other species, tobacco. Some other important members of Solanaceae include a number of ornamental plants such as Petunia, Browallia, and Lycianthes, and sources of psychoactive alkaloids, Datura, Mandragora (mandrake), and Atropa belladonna (deadly nightshade). Certain species are widely known for their medicinal uses, their psychotropic effects, or for being poisonous.
Most of the economically important genera are contained in the subfamily Solanoideae, with the exceptions of tobacco (Nicotiana tabacum, Nicotianoideae) and petunia (Petunia × hybrida, Petunioideae),
Many of the Solanaceae, such as tobacco and petunia, are used as model organisms in the investigation of fundamental biological questions at the cellular, molecular, and genetic levels.
Etymology and pronunciation
The name "Solanaceae" (US English /ˌsoʊləˈneɪsi, -siˌaɪ, -siˌeɪ, -siˌi/) comes to international scientific vocabulary from New Latin, from Solanum, the type genus, + -aceae,[4] a standardized suffix for plant family names in modern taxonomy. The genus name comes from the Classical Latin word solanum, referring to nightshades (especially Solanum nigrum), "probably from sol, 'sun', + -anum, neuter of -anus."[4]
Description

Plants in the "Solanaceae" can take the form of herbs, shrubs, trees, vines and lianas, and sometimes epiphytes. They can be annuals, biennials, or perennials, upright or decumbent. Some have subterranean tubers. They do not have laticifers, nor latex, nor coloured saps. They can have a basal or terminal group of leaves or neither of these types. The leaves are generally alternate or alternate to opposed (that is, alternate at the base of the plant and opposed towards the inflorescence). The leaves can be herbaceous, leathery, or transformed into spines. The leaves are generally petiolate or subsessile, rarely sessile. They are frequently inodorous, but on occasions, they are aromatic or fetid. The foliar lamina can be either simple or compound, and the latter can be either pinnatifid or ternate. The leaves have reticulated venation and lack a basal meristem. The laminae are generally dorsiventral and lack secretory cavities. The stomata are generally confined to one of a leaf's two sides; they are rarely found on both sides.
The flowers are generally hermaphrodites, although some are monoecious, andromonoecious, or dioecious species (such as some Solanum or Symonanthus). Pollination is entomophilous. The flowers can be solitary or grouped into terminal, cymose, or axillary inflorescences. The flowers are medium-sized, fragrant (Nicotiana), fetid (Anthocercis), or inodorous. The flowers are usually actinomorphic, slightly zygomorphic, or markedly zygomorphic (for example, in flowers with a bilabial corolla in Schizanthus species). The irregularities in symmetry can be due to the androecium, to the perianth, or both at the same time. In the great majority of species, the flowers have a differentiated perianth with a calyx and corolla (with five sepals and five petals, respectively) an androecium with five stamens and two carpels forming a gynoecium with a superior ovary[5] (they are therefore referred to as pentamers and tetracyclic). The stamens are epipetalous and are typically present in multiples of four or five, most commonly four or eight. They usually have a hypogynous disk. The calyx is gamosepalous (as the sepals are joined together forming a tube), with the (4)5(6) segments equal, it has five lobes, with the lobes shorter than the tube, it is persistent and often accrescent. The corolla usually has five petals that are also joined together forming a tube. Flower shapes are typically rotate (wheel-shaped, spreading in one plane, with a short tube) or tubular (elongated cylindrical tube), campanulated or funnel-shaped.
The androecium has (2)(4)5(6) free stamens within it, oppositsepals (that is, they alternate with the petals), they are usually fertile or, in some cases (for example in Salpiglossideae) they have staminodes. In the latter case, there is usually either one staminode (Salpiglossis) or three (Schizanthus). The anthers touch on their upper end forming a ring, or they are completely free, dorsifixed, or basifixed with poricide dehiscence or through small longitudinal cracks. The stamen’s filament can be filliform or flat. The stamens can be inserted inside the coralline tube or exserted. The plants demonstrate simultaneous microsporogenesis, the microspores are tetrad, tetrahedral, or isobilateral. The pollen grains are bicellular at the moment of dehiscence, usually open and angular.
The gynoecium is bicarpelar (rarely three- or five-locular) with a superior ovary and two locules, which may be secondarily divided by false septa, as is the case for Nicandreae and Datureae. The gynoecium is located in an oblique position relative to the flower’s median plane. They have one style and one stigma; the latter is simple or bilobate. Each locule has one to 50 ovules that are anatropous or hemianatropous with axillar placentation. The development of the embryo sack can be the same as for Polygonum or Allium species. The embryo sack’s nuclear poles become fused before fertilization. The three antipodes are usually ephemeral or persistent as in the case of Atropa. The fruit can be a berry as in the case of the tomato or wolfberry a dehiscent capsule as in Datura, or a drupe. The fruit has axial placentation. The capsules are normally septicidal or rarely loculicidal or valvate. The seeds are usually endospermic, oily (rarely starchy), and without obvious hairs. The seeds of most Solanaceae are round and flat, about 2–4 mm (0.079–0.157 in) in diameter. The embryo can be straight or curved, and has two cotyledons. Most species in the Solanaceae have 2n=24 chromosomes,[6] but the number may be a higher multiple of 12 due to polyploidy. Wild potatoes, of which there are about 200, are predominantly diploid (2 × 12 = 24 chromosomes), but triploid (3 × 12 = 36 chromosomes), tetraploid (4 × 12 = 48 chromosomes), pentaploid (5 × 12 = 60) and even hexaploid (6 × 12 = 72 chromosome) species or populations exist. The cultivated species Solanum tuberosum has 4 × 12 = 48 chromosomes. Some Capsicum species have 2 × 12 = 24 chromosomes, while others have 26 chromosomes.
Diversity of characteristics
Despite the previous description, the Solanaceae exhibit a large morphological variability, even in their reproductive characteristics. Examples of this diversity include:[7][8]
- The number of carpels that form the gynoecium
In general, the Solanaceae have a gynoecium (the female part of the flower) formed of two carpels. However, Melananthus has a monocarpelar gynoecium, there are three or four carpels in Capsicum, three to five in Nicandra, some species of Jaborosa and Trianaea and four carpels in Iochroma umbellatum.
- The number of locules in the ovary
The number of locules in the ovary is usually the same as the number of carpels. However, some species occur in which the numbers are not the same due to the existence of false septa (internal walls that subdivide each locule), such as in Datura and some members of the Lycieae (the genera Grabowskia and Vassobia).
- Type of ovules and their number
The ovules are generally inverted, folded sharply backwards (anatropous), but some genera have ovules that are rotated at right angles to their stalk (campilotropous) as in Phrodus, Grabowskia or Vassobia), or are partially inverted (hemitropous as in Cestrum, Capsicum, Schizanthus and Lycium). The number of ovules per locule also varies from a few (two pairs in each locule in Grabowskia, one pair in each locule in Lycium) and very occasionally only one ovule is in each locule as for example in Melananthus.
- The type of fruit
The fruits of the great majority of the Solanaceae are berries or capsules (including pyxidia) and less often drupes. Berries are common in the subfamilies Cestroideae, Solanoideae (with the exception of Datura, Oryctus, Grabowskia and the tribe Hyoscyameae) and the tribe Juanulloideae (with the exception of Markea). Capsules are characteristic of the subfamilies Cestroideae (with the exception of Cestrum) and Schizanthoideae, the tribes Salpiglossoideae and Anthocercidoideae, and the genus Datura. The tribe Hyoscyameae has pyxidia. Drupes are typical of the Lycieae tribe and in Iochrominae.
Alkaloids
Alkaloids are nitrogenous organic substances produced by plants as a secondary metabolite and which have an intense physiological action on animals even at low doses. Solanaceae are known for having a diverse range of alkaloids. To humans, these alkaloids can be desirable, toxic, or both. The tropanes are the most well-known of the alkaloids found in the Solanaceae. The plants that contain these substances have been used for centuries as poisons. However, despite being recognized as poisons, many of these substances have invaluable pharmaceutical properties. Many species contain a variety of alkaloids that can be more or less active or poisonous, such as scopolamine, atropine, hyoscyamine, and nicotine. They are found in plants such as the henbane (Hyoscyamus albus), belladonna (Atropa belladonna), datura or jimson (Datura stramonium), mandrake (Mandragora autumnalis), tobacco, and others. Some of the main types of alkaloids are:

- Solanine: A toxic glycoalkaloid with a bitter taste, it has the formula C45H73NO15. It is formed by the alkaloid solanidine with a carbohydrate side chain. It is found in leaves, fruit, and tubers of various Solanaceae such as the potato and tomato. Its production is thought to be an adaptive defence strategy against herbivores. Substance intoxication from solanine is characterized by gastrointestinal disorders (diarrhoea, vomiting, abdominal pain) and neurological disorders (hallucinations and headache). The median lethal dose is between 2 and 5 mg per kg of body weight. Symptoms become manifest 8 to 12 hr after ingestion. The amount of these glycoalkaloids in potatoes, for example, varies significantly depending of environmental conditions during their cultivation, the length of storage, and the variety. The average glycoalkaloid concentration is 0.075 mg/g of potato.[9] Solanine has occasionally been responsible for poisonings in people who ate berries from species such as Solanum nigrum or Solanum dulcamara, or green potatoes.[10][11]

- Tropanes: The term "tropane" comes from a genus in which they are found, Atropa (the belladonna genus). Atropa is named after the Greek Fate, Atropos, who cut the thread of life. This nomenclature reflects its toxicity and lethality. They are bicyclic organic nitrogen compounds (IUPAC nomenclature: 8-Methyl-8-azabicyclo[3.2.1]octane), with the chemical formula of C8H15N. These alkaloids include, among others, atropine, cocaine, scopolamine, and hyoscyamine. They are found in various species, such as mandrake (Mandragora autumnalis), black henbane or stinking nightshade (Hyoscyamus niger), belladonna (Atropa belladonna) the stramonium (Datura stramonium) and Brugmansia species, as well as many others in the Solanaceae family.[12] Pharmacologically, they are the most powerful known anticholinergics in existence, meaning they inhibit the neurological signals transmitted by the endogenous neurotransmitter, acetylcholine. More commonly, they can halt many types of allergic reactions. Symptoms of overdose may include dry mouth, dilated pupils, ataxia, urinary retention, hallucinations, convulsions, coma, and death. Atropine, a commonly used ophthalmological agent, dilates the pupils and thus facilitates examination of the interior of the eye. In fact, juice from the berries of A. belladonna were used by Italian courtesans during the Renaissance to exaggerate the size of their eyes by causing the dilation of their pupils. Despite the extreme toxicity of the tropanes, they are useful drugs when administered in extremely small dosages. They can reverse cholinergic poisoning, which can be caused by overexposure to organophosphate insecticides and chemical warfare agents such as sarin and VX. Scopolamine (found in Hyoscyamus muticus and Scopolia atropioides), is used as an antiemetic against motion sickness or for people suffering from nausea as a result of receiving chemotherapy.[13][14] Scopolamine and hyoscyamine are the most widely used tropane alkaloids in pharmacology and medicine due to their effects on the parasympathetic nervous system. Atropine has a stimulant effect on the central nervous system and heart, whereas scopolamine has a sedative effect. These alkaloids cannot be substituted by any other class of compounds, so they are still in demand. This is one of the reasons for the development of an active field of research into the metabolism of the alkaloids, the enzymes involved, and the genes that produce them. Hyoscyamine 6-β hydroxylase, for example, catalyses the hydroxylation of hyoscyamine that leads to the production of scopolamine at the end of the tropane’s biosynthetic pathway. This enzyme has been isolated and the corresponding gene cloned from three species: H. niger, A. belladonna and B. candida.[15][16][17]
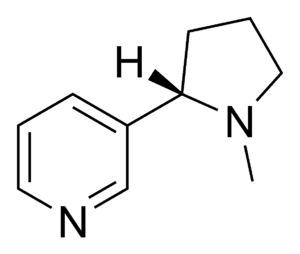
- Nicotine: Nicotine (IUPAC nomenclature (S)-3-(1-methylpyrrolidin-2-il) pyridine) is a pyrrolidine alkaloid produced in large quantities in the tobacco plant (Nicotiana tabacum), but is also found in lower concentrations in other species such as the potato, tomato, and pepper. Its function in a plant is to act as a defence against herbivores, as it is an excellent neurotoxin, in particular against insects. In fact, nicotine has been used for many years as an insecticide, although its use is currently being replaced by synthetic molecules derived from its structure. At low concentrations, nicotine acts as a stimulant in mammals, which causes the dependency in smokers. Like the tropanes, it acts on cholinergic neurons, but with the opposite effect (it is an agonist as opposed to an antagonist). It has a higher specificity for nicotinic acetylcholine receptors than other ACh proteins.

- Capsaicin: Capsaicin (IUPAC nomenclature 8-methyl-N-vanillyl-trans-6-nonenamide) is structurally different from nicotine and the tropanes. It is found in species of the genus Capsicum, which includes chilies and habaneros and it is the active ingredient that determines the Scoville rating of these spices. The compound is not noticeably toxic to humans. However, it stimulates specific pain receptors in the majority of mammals, specifically those related to the perception of heat in the oral mucosa and other epithelial tissues. When capsaicin comes into contact with these mucosae, it causes a burning sensation little different from a burn caused by fire. Capsaicin affects only mammals, not birds. Pepper seeds can survive the digestive tracts of birds; their fruit becomes brightly coloured once its seeds are mature enough to germinate, thereby attracting the attention of birds that then distribute the seeds. Capsaicin extract is used to make pepper spray, a useful deterrent against aggressive mammals.
Distribution

Even though members of the Solanaceae are found on all continents except Antarctica, the greatest variety of species are found in Central America and South America. Centers of diversity also occur in Australia and Africa. Solanaceae occupy a great number of different ecosystems, from deserts to rainforests, and are often found in the secondary vegetation that colonizes disturbed areas. In general, plants in this family are of tropical and temperate distribution.
Taxonomy
The following taxonomic synopsis of the solanaceas, including subfamilies, tribes and genera, is based on the most recent molecular phylogenetics studies of the family:[2][3][18][19]

Cestroideae (Browallioideae)



This subfamily is characterised by the presence of pericyclic fibres, an androecium with four or five stamens, frequently didynamous. The basic chromosome numbers are highly variable, from x=7 to x=13. The subfamily consists of eight genera (divided into three tribes) and about 195 species distributed throughout the Americas. The Cestrum genus is the most important, as it contains 175 of the 195 species in the subfamily. The Cestreae tribe is unusual because it includes taxa with long chromosomes (from 7.21 to 11.511 µm in length), when the rest of the family generally possesses short chromosomes (for example between 1.5 and 3.52 µm in the Nicotianoideae)
- Browallieae Hunz.
- Browallia L., genus with six species distributed throughout the neotropic ecozone to Arizona in the United States
- Streptosolen Miers, monotypic genus native to the Andes
- Cestreae tribe Don, three genera of woody plants, generally shrubs
- Salpiglossideae tribe (Benth.) Hunz.
- Reyesia Gay, four species, distributed throughout Argentina and Chile
- Salpiglossis Ruiz & Pav., two species originating from southern South America
Goetzeoideae
This subfamily is characterized by the presence of drupes as fruit and seeds with curved embryos and large fleshy cotyledons. The basic chromosome number is x=13. It includes four genera and five species distributed throughout the Greater Antilles. Some authors suggest their molecular data indicate the monotypic genera Tsoala Bosser & D'Arcy should be included in this subfamily, endemic to Madagascar, and Metternichia to the southeast of Brazil. Goetzeaceae Airy Shaw is considered as a synonym of this subfamily.[20]
- Coeloneurum Radlk., monotypic genus endemic to Hispaniola
- Espadaea Rchb., monotypic, from Cuba
- Goetzea Wydler, includes two species from the Antilles
- Henoonia Griseb., monotypic, originating in Cuba
Petunioideae

Molecular phylogenetics indicates that Petunioideae is the sister clade of the subfamilies with chromosome number x=12 (Solanoideae and Nicotianoideae). They contain calistegins, alkaloids similar to the tropanes. The androecium is formed of four stamens (rarely five), usually with two different lengths. The basic chromosome number of this subfamily can be x=7, 8, 9 or 11. It consists of 13 genera and some 160 species distributed throughout Central and South America. Molecular data suggest the genera originated in Patagonia. Benthamiella, Combera, and Pantacantha form a clade that can be categorized as a tribe (Benthamielleae) that should be in the subfamily Goetzeoideae.
- Benthamiella Speg., 12 species native to Patagonia
- Bouchetia Dunal, three neotropical species
- Brunfelsia L., around 45 species from the neotropics
- Combera Sandw., two species from Patagonia
- Fabiana Ruiz & Pav., 15 species native to the Andes
- Hunzikeria D'Arcy, three species from the southwest United States and Mexico
- LatuaPhil., one species from the south of Chile
- Leptoglossis Benth., seven species from western South America
- Nierembergia Ruiz & Pav., 21 species from South America
- Pantacantha Speg., monospecific genus from Patagonia
- Calibrachoa Cerv. ex La Llave & Lex. consists of 32 species from the neotropics. The morphological data suggest this genus should be included within the Petunia. However, the molecular and cytogenetic data indicate both should be kept separate. In fact, Calibrachoa has a basic chromosome number x=9, while that of Petunia is x=7.[21][22]
- Petunia (Juss.) Wijsman, 18 species from South America
- Plowmania Hunz. & Subils, monotypic genus from Mexico and Guatemala
Schizanthoideae

The Schizanthoideae include annual and biennial plants with tropane alkaloids, without pericyclic fibres, with characteristic hair and pollen grains. The flowers are zygomorphic. The androecium has two stamens and three stamenodes, anther dehiscence is explosive. The embryo is curved. The basic chromosome number is x=10. Schizanthus is a somewhat atypical genus among the Solanaceae due to its strongly zygomorphic flowers and basic chromosome number. Morphological and molecular data suggest Schizanthus is a sister genus to the other Solanaceae and diverged early from the rest, probably in the late Cretaceous or in the early Cenozoic, 50 million years ago.[18][19] The great diversity of flower types within Schizanthus has been the product of the species’ adaptation to the different types of pollinators that existed in the Mediterranean, high alpine, and desert ecosystems then present in Chile and adjacent areas of Argentina.[23]
- Schizanthus Ruiz & Pav., 12 species originating from Chile.
Schwenckioideae
Annual plants with pericyclic fibres, their flowers are zygomorphic, the androecium has four didynamous stamens or three stamenodes; the embryo is straight and short. The basic chromosome number is x=12. It includes four genera and some 30 species distributed throughout South America.
- Heteranthia Nees & Mart., one species from Brazil
- Melananthus Walp., five species from Brazil, Cuba, and Guatemala
- Protoschwenckia Soler , monotypic genus from Bolivia and Brazil, some molecular phylogenetic studies have suggested this genus has an uncertain taxonomic position within the subfamily
- Schwenckia L., 22 species distributed throughout the neotropical regions of America
Nicotianoideae

- Anthocercideae G. Don: This tribe, endemic to Australia, contains 31 species in seven genera. Molecular phylogenetic studies of the tribe indicate it is the sister of Nicotiana, and the genera Anthocercis, Anthotroche, Grammosolen, and Symonanthus are monophyletic. Some characteristics are also thought to be derived from within the tribe, such as the unilocular stamens with semicircular opercula, bracteolate flowers, and berries as fruit.[24]
- Anthocercis Labill., 10 species, Australia
- Anthotroche Endl., four species, Australia
- Crenidium Haegi, monotypic genus, Australia
- Cyphanthera Miers, 9 species, Australia
- Duboisia R.Br., four species, Australia
- Gramnosolen Haegi, two species, Australia
- Symonanthus Haegi, two species, Australia
- Nicotianeae tribe Dum.
- Nicotiana L., genus widely distributed, with 52 American species, 23 Australian, and one African
Solanoideae











- Capsiceae Dumort
- Capsicum L. includes 40 accepted neotropical species[25]
- Lycianthes (Dunal) Hassler, some 200 species distributed throughout America and Asia
- Datureae G. Don, two genera are perfectly differentiated at both the morphological and molecular levels, Brugmansia includes tree species, while Datura contains herbs or shrubs, the latter genus can be divided into three sections: Stramonium, Dutra and Ceratocaulis.[26]
- Brugmansia Persoon, six species from the Andes
- Datura L., 12 neotropical species
- Hyoscyameae Endl.
- Anisodus Link, four species from China, India and the Himalayas
- Atropa L., four Euro-Asiatic species[25]
- Atropanthe Pascher, monotypic genus from China
- Hyoscyamus L., 10 accepted species[25] distributed from the Mediterranean to China
- Physochlaina G. Don, 6 accepted Euro-Asiatic species[25]
- Przewalskia Maxim., one species from China
- Scopolia Jacq., disjointed distribution with one European species and another from Japan
- Jaboroseae Miers
- Jaborosa Juss., genus that includes 23 species from South America.
- Solandreae Miers
- Subtribe Juanulloinae consists 10 genera of trees and epiphytic shrubs with a neotropical distribution .[27] Some of these genera (Dyssochroma, Merinthopodium and Trianaea) show a clear dependency on various species of bats both for pollination and dispersion of seeds.[28]
- Dyssochroma Miers, two species from the south of Brazil
- Ectozoma Miers
- Hawkesiophyton Hunz.
- Juanulloa Ruiz & Pav., 11 species from South and Central America
- Markea Rich., 9 species from South and Central America
- Merinthopodium J. Donn. Sm. three species originating from South America
- Rahowardiana D' Arcy
- Schultesianthus Hunz., eight neotropical species
- Trianaea Planch. & Linden, six South American species
- Subtribe Solandrinae, a monotypical subtribe, differs from Juanulloinae in that its embryos have incumbent cotyledons and semi-inferior ovaries.[27]
- Solandra Sw., 10 species from the neotropical regions of America
- Subtribe Juanulloinae consists 10 genera of trees and epiphytic shrubs with a neotropical distribution .[27] Some of these genera (Dyssochroma, Merinthopodium and Trianaea) show a clear dependency on various species of bats both for pollination and dispersion of seeds.[28]
- Lycieae Hunz. has three genera of woody plants, which grow in arid or semiarid climates. The cosmopolitan genus Lycium is the oldest in the tribe and it has the greatest morphological variability.[29] Molecular phylogenetic studies suggest both Grabowskia and Phrodus should be included in the Lycium,[30] and this genus, along with Nolana and Sclerophylax, form a clade (Lyciina), which currently lacks a taxonomic category.[20] The red fleshy berries dispersed by birds are the main type of fruit in Lycium. The different types of fruit in this genus have evolved from the type of berry just mentioned to a drupe with a reduced number of seeds.[31]
- Grabowskia Schltdl., three species from South America
- Lycium L., 83 cosmopolitan species
- Phrodus Miers, two species endemic to the north of Chile
- Mandragoreae (Wettst.) Hunz. & Barboza tribe does not have a defined systematic position according to molecular phylogenetic studies.[20]
- Mandragora L., two species from Eurasia
- Nicandreae Wettst. is a tribe with two South American genera. Molecular phylogenetic studies indicate the genera are not interrelated nor are they related with other genera of the family, so their taxonomic position is uncertain.[20]
- Exodeconus Raf., six species from western South America
- Nicandra Adans, one species distributed throughout neotropical regions
- Nolaneae Rchb. are mostly herbs and small shrubs with succulent leaves, they have very beautiful flowers that range from white to various shades of blue, their fruit is schizocarpal, giving rise to various nuts.
- Nolana L., 89 species distributed throughout western South America
- Physaleae Miers, is a large tribe that is the sister of Capsiceae.
- Subtribe Iochrominae (Miers) Hunz., a clade within the Physaleae tribe. contains 37 species, mainly distributed in the Andes, assigned to six genera. The members of this subtribe are characterized by being woody shrubs or small trees with attractive tubular or rotated flowers. They also possess great floral diversity, containing every type is present in the family. Their flowers can be red, orange, yellow, green, blue, purple, or white. The corolla can be tubular to rotated, with a variation of up to eight times in the length of the tube between the various species.[32]
- Acnistus Schott, one species distributed throughout the neotropics
- Dunalia Kunth., five species from the Andes
- Iochroma Benth., 24 species from the Andes
- Saracha Ruiz & Pav., two species from the Andes.
- Vassobia Rusby, two South American species
- Eriolarynx Hunz., three species from Argentina and Bolivia
- Physalinae (Miers) Hunz. , a monophyletic subtribe, contains 10 genera and includes herbs or woody shrubs with yellow, white, or purple solitary axillary flowers pollinated by bees. Once pollination occurs, the corolla falls and the calyx expands until it entirely covers the boll that is developing (the calyx is called accrescent). In many species, the calyx turns yellow or orange on maturity. The berries contain many greenish to yellow-orange seeds, often with red or purple highlights.[33]
- Brachistus Miers, three species from Mexico and Central America
- Chamaesaracha (A.Gray) Benth. & Hook., has 10 species from Mexico and Central America.
- Leucophysalis Rydberg, includes 3 species from the south west of the United States and Mexico.
- Margaranthus Schlecht., with 1 species from Mexico.
- Oryctes S. Watson, monotypic genus from the south west of the United States.
- Quincula Raf. with just 1 species from the south west of the United States and from Mexico.
- Physalis L., the largest genus of the subtribe, with 85 species distributed through the tropical regions of the Americas and with 1 species in China.
- Witheringia L' Heritier, genus with 15 species from neotropical regions.
- Tzeltalia, genus segregated from Physalis, with 2 species distributed throughout Mexico and Guatemala.
- Darcyanthus, genus with just 1 species originating in Bolivia and Peru.
- Subtribe Salpichroinae, this is a subtribe of Physaleae that includes 16 American species distributed in 1 genera:
- Nectouxia Kunth., monotypic genus that is endemic to Mexico.
- Salpichroa Miers, genus with 15 species from the Andes and other regions of South America.
- Subtribe Withaninae, is a subtribe of Physaleae with a broad distribution, including 9 genera:
- Archiphysalis Kuang, with 3 species from China and Japan.
- Athenaea Sendtn., which includes 7 species from Brazil.
- Aureliana Sendtn., with 5 species from South America.
- Melissia Hook. f., monotypic genus from Santa Elena with the common name St. Elena boxwood.
- Physalisastrum Makino, with 9 Asiatic species.
- Tubocapsicum (Wettst.) Makino, with just one species endemic to China.
- Withania Pauq., with 10 species native to the Canary Islands, Africa and Nepal.
- Cuatresia Hunz., with 11 neotropical species. Molecular studies indicate that this genus, along with Deprea and Larnax has an uncertain taxonomic position.[20]
- Deprea Raf., with 6 neotropical species.
- Larnax Miers, many taxonomists consider it to be a synonym for Deprea, contains 22 species native to the Andes.
- Subtribe Iochrominae (Miers) Hunz., a clade within the Physaleae tribe. contains 37 species, mainly distributed in the Andes, assigned to six genera. The members of this subtribe are characterized by being woody shrubs or small trees with attractive tubular or rotated flowers. They also possess great floral diversity, containing every type is present in the family. Their flowers can be red, orange, yellow, green, blue, purple, or white. The corolla can be tubular to rotated, with a variation of up to eight times in the length of the tube between the various species.[32]
- Tribe Solaneae. The genera Cyphomandra Sendtn., Discopodium Hochst. , Normania Lowe, Triguera Cav. and Lycopersicum Mill have been transferred to Solanum. The subtribe is therefore composed of two genera:[20]
- Genera with doubtful taxonomic positions (Incertae sedis)
The following genera have still not been placed in any of the recognized subfamilies within the solanaceas.
- Duckeodendron Kuhlmannb, monotypic genus from the Amazon rainforest.
- Parabouchetia Baillon, poorly-known,monotypic genus from Brazil.
- Pauia Deb. & Dutta, monotypic genus from Assam and Arunachal Pradesh in N.E.India
Genera and distribution of species

The Solanaceae contain 98 genera and some 2,700 species. Despite this immense richness of species, they are not uniformly distributed between the genera. The eight most important genera contain more than 60% of the species, as shown in the table below. Solanum – the genus that typifies the family - includes nearly 50% of the total species of the solanaceas.
| Genera | Approximate number of species |
|---|---|
| Solanum | 1,330 |
| Lycianthes | 200 |
| Cestrum | 150 |
| Nolana | 89 |
| Physalis | 85 |
| Lycium | 85 |
| Nicotiana | 76 |
| Brunfelsia | 45 |
| Estimated number of species in the family | 2,700 |
Economic importance


The solanaceas include such important food species as the potato (Solanum tuberosum), the tomato (Solanum lycopersicum), the pepper (Capsicum annuum) and the aubergine or egg plant (Solanum melongena). Nicotiana tabacum, originally from South America, is now cultivated throughout the world to produce tobacco. Many solanaceas are important weeds in various parts of the world. Their importance lies in the fact that they can host pathogens or diseases of the cultivated plants, therefore their presence increases the loss of yield or the quality of the harvested product. An example of this can be seen with Acnistus arborescens and Browalia americana that host thrips, which cause damage to associated cultivated plants,[34] and certain species of Datura that play host to various types of virus that are later transmitted to cultivated solanaceas.[35] Some species of weeds such as, for example Solanum mauritianum in South Africa represent such serious ecological and economic problems that studies are being carried out with the objective of developing a biological control through the use of insects.[36]
Various solanaceas species are grown as ornamental trees or shrubs.[37] Examples include Brugmansia x candida ("Angel’s Trumpet") grown for its large pendulous trumpet-shaped flowers, or Brunfelsia latifolia, whose flowers are very fragrant and change colour from violet to white over a period of 3 days. Other shrub species that are grown for their attractive flowers are Lycianthes rantonnetii (Blue Potato Bush or Paraguay Nightshade) with violet-blue flowers and Nicotiana glauca ("Tree Tobacco") Other solanacea species and genera that are grown as ornamentals are the petunia (Petunia × hybrida), Lycium, Solanum, Cestrum, Calibrachoa × hybrida and Solandra. There is even a hybrid between Petunia and Calibrachoa (which constitutes a new nothogenus called × Petchoa G. Boker & J. Shaw) that is being sold as an ornamental.[38][39] Many other species, in particular those that produce alkaloids, are used in pharmacology and medicine (Nicotiana, Hyoscyamus, and Datura).
Solanaceas and the genome
Many of the species belonging to this family, among them tobacco and the tomato, are model organisms that are used for research into fundamental biological questions. One of the aspects of the solanaceas’ genomics is an international project that is trying to understand how the same collection of genes and proteins can give rise to a group of organisms that are so morphologically and ecologically different. The first objective of this project was to sequence the genome of the tomato. In order to achieve this each of the 12 chromosomes of the tomato’s haploid genome was assigned to different sequencing centres in different countries. So chromosomes 1 and 10 were sequenced in the United States, 3 and 11 in China, 2 in Korea, 4 in Britain, 5 in India, 7 in France, 8 in Japan, 9 in Spain and 12 in Italy. The sequencing of the mitochondrial genome was carried out in Argentina and the chloroplast genome was sequenced in the European Union.[40][41]
See also
- List of plants poisonous to equines
- Solanaceae Resources on the Web
- Jäpelt RB, Jakobsen J (2013) Vitamin D in plants: a review of occurrence, analysis, and biosynthesis. Front Plant Sci 4, No. 136 -- Note the reference to higher cholesterol levels (and consequent Vitamin D3 levels) in family Solanaceae
References
- ↑ "Solanaceae Juss., nom. cons.". Germplasm Resources Information Network. United States Department of Agriculture. 2007-04-12. Retrieved 2009-04-16.
- 1 2 Olmstead, R. G.; Sweere, J. A.; Spangler, R. E.; Bohs, L.; Palmer, J. D. (1999). "Phylogeny and provisional classification of the Solanaceae based on chloroplast DNA" (PDF). In Nee, M.; Symon, D. E.; Lester, R. N.; Jessop, J. P. Solanaceae IV: advances in biology and utilization. The Royal Botanic Gardens. pp. 111–37.
- 1 2 Olmstead, R.G.; Bohs, L. (2007). "A Summary of molecular systematic research in Solanaceae: 1982-2006". Acta Horticulturae. 745: 255–68.
- 1 2 Merriam-Webster, Merriam-Webster's Unabridged Dictionary, Merriam-Webster.
- ↑ Yasin J. Nasir. "Solanaceae". Flora of Pakistan.
- ↑ Fujii, Kenjiro (1934). Cytologia. Botanical Institute. p. 281.
- ↑ Hunziker, A.T. 1979: South American Solanaceae: a synoptic review. In: D'ARCY, W.G., 1979: The Biology and Taxonomy of the Solanaceae. Linn. Soc. Symp. Ser. 7: p 48-85. Linnean Soc. & Academic Press; London.
- ↑ Balken, J.A. THE PLANT FAMILY SOLANACEAE: FRUITS IN SOLANACEAE
- ↑ Zeiger, E. 1998. Solanine and Chaconine. Review of Toxicological Literature. Integrated Laboratory Systems, USA.
- ↑ "Solanine poisoning". Br Med J. 2 (6203): 1458–9. 1979-12-08. doi:10.1136/bmj.2.6203.1458-a. PMC 1597169
 . PMID 526812.
. PMID 526812. - ↑ Alexander RF, Forbes GB, Hawkins ES (1948-09-11). "A Fatal Case of Solanine Poisoning". Br Med J. 2 (4575): 518. doi:10.1136/bmj.2.4575.518. PMC 2091497. PMID 18881287.
- ↑ Griffin WJ, Lin GD (March 2000). "Chemotaxonomy and geographical distribution of tropane alkaloids". Phytochemistry. 53 (6): 623–37. doi:10.1016/S0031-9422(99)00475-6. PMID 10746874.
- ↑ Sneden, A. The tropane alkaloids. Medicinal Chemistry and Drug Design. Virginia Commonwealth University
- ↑ Evans, W.C. 1979. Tropane alkaloids of the Solanaceae. En: HAWKES, LESTER and SHELDING (eds.). The biology and taxonomy of the Solanaceae. Linn. Soc. Symp. Ser. 7:241-254. Linnean Soc. & Academic Press., London.
- ↑ Matsuda, Jun; Okabe, Souichi; Hashimoto, Takashi; Yamada, Yasuyuki (1991). "Molecular cloning of hyoscyamine 6β-hydroxylase, a 2-oxoglutarate-dependent dioxygenase, from cultured roots of Hyoscyamus niger". The Journal of Biological Chemistry. 266 (15): 9460–4. PMID 2033047.
- ↑ Rocha, Pedro; Stenzel, Olaf; Parr, Adrian; Walton, Nicholas; Christou, Paul; Dräger, Birgit; Leech, Mark J (June 2002). "Functional expression of tropinone reductase I (trI) and hyoscyamine-6β-hydroxylase (h6h) from Hyoscyamus niger in Nicotiana tabacum". Plant Science. 162 (6): 905–13. doi:10.1016/S0168-9452(02)00033-X.
- ↑ Cardillo, Alejandra B.; Giulietti, Ana M.; Marconi, Patricia L. (June 2006). "Analysis and sequencing of h6hmRNA, last enzyme in the tropane alkaloids pathway from anthers and hairy root cultures of Brugmansia candida (Solanaceae)". Electronic Journal of Biotechnology. 9 (3). doi:10.4067/S0717-34582006000300004.
- 1 2 Olmstead, Richard G.; Palmer, Jeffrey D. (1992). "A Chloroplast DNA Phylogeny of the Solanaceae: Subfamilial Relationships and Character Evolution". Annals of the Missouri Botanical Garden. 79 (2): 346–60. doi:10.2307/2399773. JSTOR 2399773.
- 1 2 Martins, Talline R.; Barkman, Todd J. (2005). "Reconstruction of Solanaceae Phylogeny Using the Nuclear Gene SAMT". Systematic Botany. 30 (2): 435–47. doi:10.1600/0363644054223675. JSTOR 25064071.
- 1 2 3 4 5 6 Olmstead, R.G.; Bohs, L. (2007). "A Summary of molecular systematic research in Solanaceae: 1982-2006". Acta Horticulturae. 745: 255–68.
- ↑ Ando, Toshio; Kokubun, Hisashi; Watanabe, Hitoshi; Tanaka, Norio; Yukawa, Tomohisa; Hashimoto, Goro; Marchesi, Eduardo; Suárez, Enrique; Basualdo, Isabel L. (2005). "Phylogenetic Analysis of Petunia sensu Jussieu (Solanaceae) using Chloroplast DNA RFLP". Annals of Botany. 96 (2): 289–97. doi:10.1093/aob/mci177. PMC 4246877. PMID 15944177.
- ↑ Mishiba, Kei-Ichiro; Ando, Toshio; Mii, Masahiro; Watanabe, Hitoshi; Kokubun, Hisashi; Hashimoto, Goro; Marchesi, Eduardo (2000). "Nuclear DNA Content as an Index Character Discriminating Taxa in the Genus Petunia sensu Jussieu (Solanaceae)". Annals of Botany. 85 (5): 665–73. doi:10.1006/anbo.2000.1122.
- ↑ Pérez, Fernanda; Arroyo, Mary T. K.; Medel, Rodrigo; Hershkovitz, Mark A. (2006). "Ancestral reconstruction of flower morphology and pollination systems in Schizanthus (Solanaceae)". American Journal of Botany. 93 (7): 1029–38. doi:10.3732/ajb.93.7.1029. PMID 21642168.
- ↑ Garcia, Vicente F.; Olmstead, Richard G. (June 2003). "Phylogenetics of Tribe Anthocercideae (Solanaceae) Based on ndhF and trnL/F Sequence Data". Systematic Botany. 28 (3): 609–15. doi:10.1043/02-52.1 (inactive 2015-01-27). JSTOR 25063900.
- 1 2 3 4 "The Plant List, Atropa". Royal Botanic Garden, Kew.
- ↑ Mace, E. S.; Gebhardt, C. G.; Lester, R. N. (1999). "AFLP analysis of genetic relationships in the tribe Datureae (Solanaceae)". Theoretical and Applied Genetics. 99 (3–4): 634–41. doi:10.1007/s001220051278. PMID 22665199.
- 1 2 Knapp, Sandra; Persson, Viveca; Blackmore, Stephen (1997). "A Phylogenetic Conspectus of the Tribe Juanulloeae (Solanaceae)". Annals of the Missouri Botanical Garden. 84 (1): 67–89. doi:10.2307/2399954. JSTOR 2399954.
- ↑ Sazima, M.; Buzato, S; Sazima, I (2003). "Dyssochroma viridiflorum (Solanaceae): A Reproductively Bat-dependent Epiphyte from the Atlantic Rainforest in Brazil". Annals of Botany. 92 (5): 725–30. doi:10.1093/aob/mcg190. PMC 4244854. PMID 14500325.
- ↑ Bernardello, Luis M. (1987). "Comparative Floral Morphology in Lycieae (Solanaceae)". Brittonia. 39 (1): 112–29. doi:10.2307/2806983. JSTOR 2806983.
- ↑ Levin, achel A.; Mille, Jill S. (2005). "Relationships within tribe Lycieae (Solanaceae): Paraphyly of Lycium and multiple origins of gender dimorphism". American Journal of Botany. 92 (12): 2044–53. doi:10.3732/ajb.92.12.2044. JSTOR 4125537. PMID 21646122.
- ↑ Bernardello, L.; Chiang-Cabrera, F. (1998). "A cladistic study on the American species of Lycium (Solanaceae) based on morphological variation". In Fortunato, Renée H; Bacigalupo, Nélida M. Proceedings of the VI Congreso Latinoamericano de Botánica, Mar del Plata, Argentina, 2-8 October, 1994. Monographs in Systematic Botany from the Missouri Botanical Garden. Missouri Botanical Garden Press. pp. 33–46. ISBN 978-0-915279-58-6.
- ↑ Smith, Stacey DeWitt; Baum, David A. (2006). "Phylogenetics of the florally diverse Andean clade Iochrominae (Solanaceae)". American Journal of Botany. 93 (8): 1140–53. doi:10.3732/ajb.93.8.1140. JSTOR 4122802. PMID 21642180.
- ↑ Whitson, Maggie; Manos, Paul S. (2005). "Untangling Physalis (Solanaceae) from the Physaloids: A Two-Gene Phylogeny of the Physalinae". Systematic Botany. 30 (1): 216–30. doi:10.1600/0363644053661841. JSTOR 25064051.
- ↑ Masis, C. & Madrigal, R. 1994. Lista preliminar de malezas hospedantes de Thrips (Thysanoptera) que dañan al Chrysanthemum morifolium en el valle central de Costa Rica. Agronomía Costarricense 18(1): 99-101. 1994
- ↑ Ormeño, J., Sepúlveda R., Rojas, R. Malezas del género Datura como factor epidemiológico del virus del mosaico de la alfalfa (amv), virus del mosaico del pepino (cmv) y virus y de la papa (pvy) en Solanáceas cultivadas. Agricultura técnica Vol. 66, Nº. 4, 2006, 333-341 Summary in Spanish
- ↑ Pedrosa-Macedo, J., Olckers, T. & Vitorino, M. 2003. Phytophagous arthropods associated with Solanum mauritianum Scopoli (Solanaceae) in the first Plateau of Paraná, Brazil: a cooperative project on biological control of weeds between Brazil and South Africa. Neotrop. Entomol. 32: 519-522. Article in English, with a summary in Portuguese
- ↑ Arboles ornamentales cultivados en España. Solanáceas
- ↑ Shaw, J. 2007. A new hybrid genus for Calibrachoa × Petunia (Solanaceae). HANBURYANA 2: 50–51
- ↑ The Value of Growing Petchoa SuperCal®. Ornamental News Oct 25 2012
- ↑ International Tomato Sequencing Project Home
- ↑ International Solanaceae Genomics Project (SOL), Systems Approach to Diversity and Adaptation.
- D'Arcy, William G. (1986). Solanaceae. Columbia University Press. ISBN 0-231-05780-6.
- Stevens, P. F. (2001–2007). "Solanaceae". Angiosperm Phylogeny Website, version 8, June 2007. Retrieved 2007-11-04.
- Watson, L.; Dallwitz, , M. J. "Solanaceae". The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 1 June 2007. Retrieved 2007-11-04.
- Dimitri, M. 1987. Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires.
- "Solanaceae Source". Retrieved 2007-11-17.
- Hunziker, Armando T. 2001. The Genera of Solanaceae. A.R.G. Gantner Verlag K.G., Ruggell, Liechtenstein. ISBN 3-904144-77-4.
Further reading
- Hawkes, J. G.; Lester, R. N.; Skelding, A. D. (1979). The biology and taxonomy of the Solanaceae. Academic Press, London. ISBN 0-12-333150-1.
- D'Arcy, William G. (1986). Solanacea. Columbia University Press. ISBN 0-231-05780-6.
- Radford, Albert E. (1986). Fundamentals of Plant Systematics. Harper & Row, Publishers, Inc. ISBN 0-06-045305-2.
- Kubo, Ken-ichi; Paape, Timothy; Hatakeyama, Masaomi; Entani, Tetsuyuki; Takara, Akie; Kajihara, Kie; Tsukahara, Mai; Shimizu-Inatsugi, Rie; Shimizu, Kentaro K.; Takayama, Seiji (2015). "Gene duplication and genetic exchange drive the evolution of S-RNase-based self-incompatibility in Petunia". Nature Plants. doi:10.1038/nplants.2014.5. Lay summary – EurekAlert! (January 8, 2015).
External links
| Wikispecies has information related to: Solanaceae |
| Wikimedia Commons has media related to Solanaceae. |
- Sol Genomics Network
- Solanaceae Network - pictures of plants
- Solanaceae Source - A worldwide taxonomic monograph of all species in the genus Solanum.
- Solanaceae of Chile, by Chileflora
- Solanaceae in L. Watson and M.J. Dallwitz (1992 onwards). The families of flowering plants: descriptions, illustrations, identification, information retrieval. http://delta-intkey.com
- Solanaceae in USDA Plants Database.
- Family Solanaceae Flowers in Israel
- SOL Genomics Network, Universidad de Cornell
- Imagines de various species of Solanaceae
- Solanaceae de Chile, by Chileflora
- Solanaceae Source, site with abundant information regarding Solanaceas
- Chilli: La especia del Nuevo Mundo (Article in Spanish by Germán Octavio López Riquelme regarding the biology, nutrition, culture and medical aspects of Chile.