Gramicidin
 | |
| Identifiers | |
|---|---|
| Symbol | N/A |
| TCDB | 1.D.1 |
| OPM superfamily | 69 |
| OPM protein | 1grm |
| Clinical data | |
|---|---|
| ATC code | R02AB30 (WHO) |
| Identifiers | |
| Synonyms | Bacillus brevis gramicidin D |
| CAS Number |
1405-97-6 |
| DrugBank |
DB00027 |
| ChemSpider |
3076403 |
| UNII |
5IE62321P4 |
| KEGG |
D04369 |
| ChEMBL |
CHEMBL557217 |
| ECHA InfoCard | 100.014.355 |
| Chemical and physical data | |
| Formula | C99H140N20O17 |
| Molar mass | 1882.3 g/mol |
| |
| | |
Gramicidin is a heterogeneous mixture of three antibiotic compounds, gramicidins A, B and C, making up 80%, 6%, and 14%, respectively,[1] all of which are obtained from the soil bacterial species Bacillus brevis and called collectively gramicidin D. Gramicidin D contains linear pentadecapeptides, that is chains made up of 15 amino acids.[2] This is in contrast to gramicidin S, which is a cyclic peptide chain.
Gramicidin was discovered in the United States in 1939.[3]
Medical uses
Gramicidin is active against Gram-positive bacteria, except for the Gram-positive bacilli, and against select Gram-negative organisms, such as Neisseria bacteria. Its therapeutic use is limited to topical application, as it induces hemolysis in lower concentrations than bacteria cell death, so it cannot be administered internally. Since the exterior epidermis is composed of dead cells, applying it to the surface of the skin will not cause harm.
It is used primarily as a topical antibiotic and is one of the three constituents of consumer antibiotic polysporin ophthalmic solution.
History
In 1939, the French microbiologist René Dubos isolated the substance tyrothricin and later showed it was composed of two substances, gramicidin (20%) and tyrocidine (80%). These were the first antibiotics to be manufactured commercially.
Composition and structure
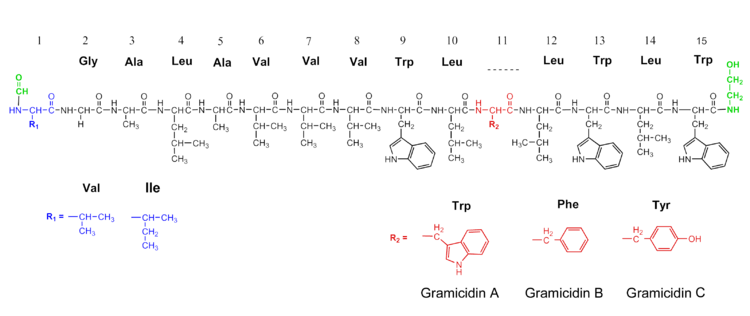
Gramicidin is a polypeptide with alternating L- and D-amino acids, sharing the general formula: formyl-L-X-Gly-L-Ala-D-Leu-L-Ala-D-Val-L-Val-D-Val-L-Trp-D-Leu-L-Y-D-Leu-L-Trp-D-Leu-L-Trp-ethanolamine
X and Y depend upon the gramicidin molecule. There exist valine and isoleucine variants of all three gramicidin species, and 'X' can be either. Y determines which is which; as Y gramicidin A contains tryptophan, B phenylalanine, and C tyrosine. Also note the alternating stereochemical configurations (in the form of D and L) of the amino acids; this is vital to the formation of the β-helix.
The chain assembles inside of the hydrophobic interior of the cellular lipid bilayer to form a β-helix. The helix itself is not long enough to span the membrane, but it dimerizes to form the elongated channel needed to span the whole membrane.
The structure of the gramicidin head-to-head dimer in micelles and lipid bilayers was determined by solution and solid-state NMR.[4] The structure was first proposed by D. W. Urry in 1971.[5] In organic solvents and crystals, this peptide forms different types of non-native double helices.
Pharmacological effect
Gramicidin's bactericidal activity is a result of increasing the permeability of the bacterial cell membrane, allowing inorganic monovalent cations (e.g. Na+) to travel through unrestricted and thereby destroying the ion gradient between the cytoplasm and the extracellular environment.[6]
That gramicidin D functions as a channel was demonstrated by Hladky and Haydon, who investigated the unitary conductance of the channel. In general, gramicidin channels are completely selective for monovalent cations and the single-channel conductances for the alkali cations are ranked in the same order as the aqueous mobilities of these ions. Divalent cations like Ca2+
block the channel by binding near its mouth so it is essentially impermeable to divalent cations and also excludes anions. Cl−
in particular is excluded from the channel because its hydration shell is thermodynamically stronger than that of most monovalent cations. The channel is permeable to most monovalent cations, which move through the channel in single file. The channel is filled with about six water molecules, almost all of which must be displaced when an ion is transported. Thus, ions moving through the gramicidin pore carry along a single file of water molecules. Such a flux of ion and water molecules is known as flux coupling. In the presence of a second type of permeable ion, the two ions couple their flux, as well. Like valinomycin and nonactin, the gramicidin channel is selective for potassium over sodium, but only slightly so. It has a permeability ratio of 2.9. Though it is impermeable to anions, conditions exist under which some anion permeation may be observed. Its ability to bind and transport cations is due to the presence of cation-binding sites, one strong and the other weak, in the channel.
References
- ↑ Bourinbaiar, A.S.; Coleman, C.F. (November 1997). "The effect of gramicidin, a topical contraceptive and antimicrobial agent with anti-HIV activity, against herpes simplex viruses type 1 and 2 in vitro". Archives of Virology. 142 (11): 2225–2235. doi:10.1007/s007050050237. PMID 9672588.
- ↑ Biopolymers (Peptide Science), vol. 51, pp. 129–144 (1999): http://www3.interscience.wiley.com/cgi-bin/abstract/62500357/ABSTRACT Burkhart, Brian M.; Gassman, R.M.; Langs, D.A.; Pangborn, W.A.; Duax, W.L.; Pletnev, V. (1999). "Gramicidin D conformation, dynamics and membrane ion transport". Biopolymers. 51 (2): 129–44. doi:10.1002/(SICI)1097-0282(1999)51:2<129::AID-BIP3>3.0.CO;2-Y. PMID 10397797.
- ↑ Levy, Stuart B. (2002). The antibiotic paradox: how the misuse of antibiotics destroys their curative powers (2 ed.). Cambridge, Mass.: Perseus Publ. p. 51. ISBN 9780738204406.
- ↑ Ketchem, Randal R.; Hu, Weidong; Cross, Timothy A. (1993). "High-Resolution Conformation of Gramicidin A in a Lipid Bilayer by Solid-State NMR". Science. 261: 1457–1460. doi:10.1126/science.7690158.
- ↑ Urry DW (1971). "The Gramicidin A Transmembrane Channel: A Proposed π(L,D) Helix". Proceedings of the National Academy of Sciences of the United States of America. 68 (3): 672–676. doi:10.1073/pnas.68.3.672. PMC 389014
 . PMID 5276779.
. PMID 5276779. - ↑ Francisco J. Alvarez-Leefmans; Eric Delpire (10 August 2009). Physiology and Pathology of Chloride Transporters and Channels in the Nervous System: From Molecules to Diseases. Academic Press. pp. 142–146. ISBN 978-0-12-374373-2. Retrieved 3 December 2010.