Sedoheptulose-bisphosphatase
| sedoheptulose-bisphosphatase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Crystallographic structure of sedoheptulose-bisphosphatase from Toxoplasma gondii [1] | |||||||||
| Identifiers | |||||||||
| EC number | 3.1.3.37 | ||||||||
| CAS number | 9055-32-7 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / EGO | ||||||||
| |||||||||
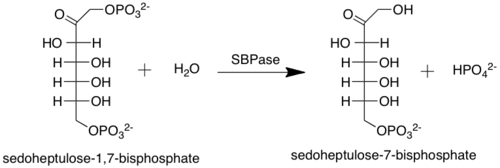
Sedoheptulose-bisphosphatase (also sedoheptulose-1,7-bisphosphatase or SBPase) (EC 3.1.3.37) is an enzyme that catalyzes the removal of a phosphate group from sedoheptulose 1,7-bisphosphate to produce sedoheptulose 7-phosphate. SBPase is an example of a phosphatase, or, more generally, a hydrolase. This enzyme participates in the Calvin cycle.
Structure
SBPase is a homodimeric protein, meaning that it is made up of two identical subunits.[2] The size of this protein varies between species, but is about 92,000 Da (two 46,000 Da subunits) in cucumber plant leaves.[3] The key functional domain controlling SBPase function involves a disulfide bond between two cysteine residues.[4] Additionally, SBPase requires the presence of magnesium (Mg2+) to be functionally active.[5] SBPase is bound to the stroma-facing side of the thylakoid membrane in the chloroplast in a plant. Some studies have suggested the SBPase may be part of a large (900 kDa) multi-enzyme complex along with a number of other photosynthetic enzymes.[6]
Regulation
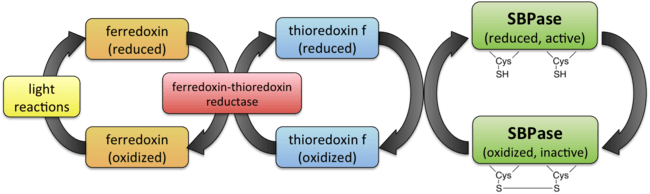
SBPase is involved in the regeneration of 5-carbon sugars during the Calvin cycle. Although SBPase has not been emphasized as an important control point in the Calvin cycle historically, it plays a large part in controlling the flux of carbon through the Calvin cycle.[7] Additionally, SBPase activity has been found to have a strong correlation with the amount of photosynthetic carbon fixation.[8] Like many Calvin cycle enzymes, SBPase is activated in the presence of light through a ferredoxin/thioredoxin system.[9] In the light reactions of photosynthesis, light energy powers the transport of electrons to eventually reduce ferredoxin. The enzyme ferredoxin-thioredoxin reductase uses reduced ferredoxin to reduce thioredoxin from the disulfide form to the dithiol. Finally, the reduced thioredoxin is used to reduced a cysteine-cysteine disulfide bond in SBPase to a dithiol, which converts the SBPase into its active form.[5]
SBPase has additional levels of regulation beyond the ferredoxin/thioredoxin system. Mg2+ concentration has a significant impact on the activity of SBPase and the rate of the reactions it catalyzes.[10] SBPase is inhibited by acidic conditions (low pH). This is a large contributor to the overall inhibition of carbon fixation when the pH is low inside the stroma of the chloroplast.[11] Finally, SBPase is subject to negative feedback regulation by sedoheptulose-7-phosphate and inorganic phosphate, the products of the reaction it catalyzes.[12]
Evolutionary Origin
SBPase and FBPase (fructose-1,6-bisphosphatase) are both phosphatases that catalyze similar during the Calvin cycle. The genes for SBPase and FBPase are related. Both genes are found in the nucleus in plants, and have bacterial ancestry.[13] SBPase is found across many species. In addition to being universally present in photosynthetic organism, SBPase is found in a number of evolutionarily-related, non-photosynthetic microorganisms. SBPase likely originated in red algae.[14]
References
- ↑ Minasov, G.; Ruan, J.; Wawrzak, Z.; Halavaty, A.; Shuvalova, L.; Harb, O. S.; Ngo, H.; Anderson, W. F. (2013). "1.85 Angstrom Crystal Structure of Putative Sedoheptulose-1,7 bisphosphatase from Toxoplasma gondii". doi:10.2210/pdb4ir8/pdb.
- ↑ Hino, M.; Nagatsu, T.; Kakumu, S.; Okuyama, S.; Yoshii, Y.; Nagatsu, I. (1975). "Glycylprolyl beta-naphthylamidase activity in human serum". Clinica chimica acta; international journal of clinical chemistry. 62 (1): 5–11. doi:10.1016/0009-8981(75)90273-9. PMID 1149281.
- ↑ Wang, M.; Bi, H.; Liu, P.; Ai, X. (2011). "Molecular cloning and expression analysis of the gene encoding sedoheptulose-1, 7-bisphosphatase from Cucumis sativus". Scientia Horticulturae. 129 (3): 414–420. doi:10.1016/j.scienta.2011.04.010.
- ↑ Anderson, L. E.; Huppe, H. C.; Li, A. D.; Stevens, F. J. (1996). "Identification of a potential redox-sensitive interdomain disulfide in the sedoheptulose bisphosphatase of Chlamydomonas reinhardtii". The Plant journal : for cell and molecular biology. 10 (3): 553–560. doi:10.1046/j.1365-313X.1996.10030553.x. PMID 8811868.
- 1 2 Nakamura, Y.; Tada, T.; Wada, K.; Kinoshita, T.; Tamoi, M.; Shigeoka, S.; Nishimura, K. (2001). "Purification, crystallization and preliminary X-ray diffraction analysis of the fructose-1,6-/sedoheptulose-1,7-bisphosphatase of Synechococcus PCC 7942". Acta Crystallographica Section D. 57 (Pt 3): 454–456. doi:10.1107/S0907444901002177. PMID 11223530.
- ↑ Ellis K.H; Arkona C.; Mantbuffel R.; Adler K. (June 1993). "Calvin cycle multienzyme complexes are bound to chloroplast thylakoid membranes of higher plants in situ". Proceedings of the National Academy of Sciences USA. 90 (12): 5514–5518. doi:10.1073/pnas.90.12.5514.
- 1 2 Raines, C. A.; Lloyd, J. C.; Dyer, T. A. (1999). "New insights into the structure and function of sedoheptulose-1,7-bisphosphatase; an important but neglected Calvin cycle enzyme". Journal of Experimental Botany. 50 (330): 1–8. doi:10.1093/jxb/50.330.1.
- ↑ Olçer, H.; Lloyd, J. C.; Raines, C. A. (2001). "Photosynthetic capacity is differentially affected by reductions in sedoheptulose-1,7-bisphosphatase activity during leaf development in transgenic tobacco plants". Plant Physiology. 125 (2): 982–989. doi:10.1104/pp.125.2.982. PMC 64898
 . PMID 11161054.
. PMID 11161054. - ↑ Breazeale VD, Buchanan BB, Wolosiuk RA (May 1978). "Chloroplast Sedoheptulose 1,7 Bisphosphatase: Evidence for Regulation by the Ferridoxin/Thioredoxin System". Zeitschrift für Naturforschung C. 33: 521–528.
- ↑ Woodrow, I. E.; Walker, D. A. (1982). "Activation of wheat chloroplast sedoheptulose bisphosphatase: A continuous spectrophotometric assay". Archives of Biochemistry and Biophysics. 216 (2): 416–422. doi:10.1016/0003-9861(82)90230-2. PMID 6287934.
- ↑ Purczeld, P.; Chon, C. J.; Portis Jr, A. R.; Heldt, H. W.; Heber, U. (1978). "The mechanism of the control of carbon fixation by the pH in the chloroplast stroma. Studies with nitrite-mediated proton transfer across the envelope". Biochimica et Biophysica Acta. 501 (3): 488–498. doi:10.1016/0005-2728(78)90116-0. PMID 24470.
- ↑ Schimkat, D.; Heineke, D.; Heldt, H. (1990). "Regulation of sedoheptulose-1,7-bisphosphatase by sedoheptulose-7-phosphate and glycerate, and of fructose-1,6-bisphosphatase by glycerate in spinach chloroplasts". Planta. 181. doi:10.1007/BF00202330.
- ↑ Martin, W.; Mustafa, A. Z.; Henze, K.; Schnarrenberger, C. (1996). "Higher-plant chloroplast and cytosolic fructose-1,6-bisphosphatase isoenzymes: Origins via duplication rather than prokaryote-eukaryote divergence". Plant molecular biology. 32 (3): 485–491. doi:10.1007/BF00019100. PMID 8980497.
- ↑ Teich, R.; Zauner, S.; Baurain, D.; Brinkmann, H.; Petersen, J. R. (2007). "Origin and Distribution of Calvin Cycle Fructose and Sedoheptulose Bisphosphatases in Plantae and Complex Algae: A Single Secondary Origin of Complex Red Plastids and Subsequent Propagation via Tertiary Endosymbioses". Protist. 158 (3): 263–276. doi:10.1016/j.protis.2006.12.004. PMID 17368985.
Further reading
- Racker E (1962). "Sedoheptulose-1,7-diphosphatase from yeast". Methods Enzymol. Methods in Enzymology. 5: 270–272. doi:10.1016/S0076-6879(62)05217-9. ISBN 978-0-12-181805-0.
- Traniello S, Calcagno M, Pontremoli S (1971). "Fructose 1,6-diphosphatase and sedoheptulose 1,7-diphosphatase from Candida utilis: purification and properties". Arch. Biochem. Biophys. 146 (2): 603–10. doi:10.1016/0003-9861(71)90168-8. PMID 4329855.