Implantation (human embryo)
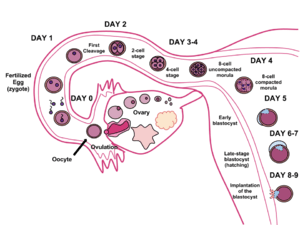
In humans (as in all other mammals, except for monotremes), implantation is the very early stage of pregnancy at which the conceptus adheres to the wall of the uterus.[1] At this stage of prenatal development, the conceptus is a blastocyst. It is by this adhesion that the fetus receives oxygen and nutrients from the mother to be able to grow.
In humans, implantation of a fertilized ovum is most likely to occur about 9 days after ovulation, ranging between 6 and 12 days.[2]
Implantation window
The reception-ready phase of the endometrium of the uterus is usually termed the "implantation window" and lasts about 4 days. The implantation window follows around 6 days after the peak in luteinizing hormone levels. With some disparity between sources, it has been stated to occur from 7 days after ovulation until 9 days after ovulation,[3] or days 6-10 postovulation.[4] On average, it occurs during the 20th to the 23rd day after the last menstrual period.[5]
The implantation window is characterized by changes to the endometrium cells, which aid in the absorption of the uterine fluid. These changes are collectively known as the plasma membrane transformation and bring the blastocyst nearer to the endometrium and immobilize it. During this stage the blastocyst can still be eliminated by being flushed out of the uterus. Scientists have hypothesized that the hormones cause a swelling that fills the flattened out uterine cavity just prior to this stage, which may also help press the blastocyst against the endometrium.[6] The implantation window may also be initiated by other preparations in the endometrium of the uterus, both structurally and in the composition of its secretions.
Adaptation of uterus
To enable implantation, the uterus goes through changes in order to be able to receive the conceptus.
Predecidualization
The endometrium increases thickness, becomes vascularized and its glands grow to be tortuous and boosted in their secretions. These changes reach their maximum about 7 days after ovulation.
Furthermore, the surface of the endometrium produces a kind of rounded cells, which cover the whole area toward the uterine cavity. This happens about 9 to 10 days after ovulation.[7] These cells are called decidual cells, which emphasises that the whole layer of them is shed off in every menstruation if no pregnancy occurs, just as leaves of deciduous trees. The uterine glands, on the other hand, decrease in activity and degenerate already 8 to 9 days[7] after ovulation in absence of pregnancy.
The decidual cells originate from the stromal cells that are always present in the endometrium. However, the decidual cells make up a new layer, the decidua. The rest of the endometrium, in addition, expresses differences between the luminal and the basal sides. The luminal cells form the zona compacta of the endometrium, in contrast to the basalolateral zona spongiosa, which consists of the rather spongy stromal cells.[7]
Decidualization
Decidualization succeeds predecidualization if pregnancy occurs. This is an expansion of it, further developing the uterine glands, the zona compacta and the epithelium of decidual cells lining it. The decidual cells become filled with lipids and glycogen and take the polyhedral shape characteristic for decidual cells.
Trigger
It is likely that the blastocyst itself makes the main contribution to this additional growing and sustaining of the decidua. An indication of this is that decidualization occurs at a higher degree in conception cycles than in nonconception cycles.[7] Furthermore, similar changes are observed when giving stimuli mimicking the natural invasion of the embryo.[7]
Parts of decidua
The decidua can be organized into separate sections, although they have the same composition.
- Decidua basalis - This is the part of the decidua which is located basalolateral to the embryo after implantation.
- Decidua capsularis - Decidua capsularis grows over the embryo on the luminal side, enclosing it into the endometrium. It surrounds the embryo together with decidua basalis.
- Decidua parietalis - All other decidua on the uterine surface belongs to decidua parietalis.
Decidua throughout pregnancy
After implantation the decidua remains, at least through the first trimester.[7] However, its most prominent time is during the early stages of pregnancy, during implantation. Its function as a surrounding tissue is replaced by the definitive placenta. However, some elements of the decidualization remain throughout pregnancy.[7]
The compacta and spongiosa layers are still observable beneath the decidua in pregnancy. The glands of the spongiosa layer continue to secrete during the first trimester, when they degenerate. However, before that disappearance, some glands secrete unequally much. This phenomenon of hypersecretion is called the Arias-Stella phenomenon,[7] after the pathologist Javier Arias-Stella.
Pinopodes
Pinopodes are small, finger-like protrusions from the endometrium. They appear between day 19 and day 21[7] of gestational age. This corresponds to a fertilization age of approximately 5 to 7 days, which corresponds well with the time of implantation. They only persist for 2 to 3 days.[7] The development of them is enhanced by progesterone but inhibited by estrogens.
Function in implantation
Pinopodes endocytose uterine fluid and macromolecules in it. By doing so, the volume of the uterus decreases, taking the walls closer to the embryoblast floating in it. Thus, the period of active pinocytes might also limit the implantation window.[7]
Function during implantation
Pinopodes continue to absorb fluid, and removes most of it during the early stages of implantation.
Adaptation of secretions
| proteins, glycoproteins and peptides
secreted by the endometrial glands[7] |
| Matrix-associated: |
| Fibronectin |
| Laminin |
| Entactin |
| Type-IV collagen |
| heparan sulfate |
| Proteoglycan |
| Integrins |
| - |
| Others: |
| Mucins |
| Prolactin |
| IGFBP-1 |
| Placental protein 14 (PP14) or glycodelin |
| Pregnancy-associated endometrial
alpha-2-globulin (alpha-2-PEG) |
| endometrial protein 15 |
| Albumin |
| Beta-Lipoprotein |
| Relaxin |
| Fibroblast growth factor 1 |
| Fibroblast growth factor 2 |
| Pregnancy-associated plasma protein A
(PAPP-A) |
| Stress response protein 27 (SRP-27) |
| CA-125 |
| Beta-endorphin |
| Leu-enkephalin |
| Diamine oxidase |
| Tissue plasminogen activator |
| Renin |
| Progesterone-dependent carbonic anhydrase |
| Lactoferrin |
Not only the lining of the uterus transforms, but in addition, the secretion from its epithelial glands changes. This change is induced by increased levels of progesterone from the corpus luteum. The target of the secretions is the embryoblast, and has several functions on it.
Nourishment
The embryoblast spends approximately 72 hours[7] in the uterine cavity before implanting. In that time, it cannot receive nourishment directly from the blood of the mother, and must rely on secreted nutrients into the uterine cavity, e.g. iron[7] and fat-soluble vitamins.[7]
Growth and implantation
In addition to nourishment, the endometrium secretes several steroid-dependent proteins,[7] important for growth and implantation. Cholesterol[7] and steroids[7] are also secreted. Implantation is further facilitated by synthesis of matrix substances, adhesion molecules and surface receptors for the matrix substances.
Mechanism
Implantation is initiated when the blastocyst comes into contact with the uterine wall.
Zona hatching
To be able to perform implantation, the blastocyst first needs to get rid of its zona pellucida. This process can be called "hatching".
Factors
Lytic factors in the uterine cavity, as well as factors from the blastocyst itself are essential for this process. Mechanisms in the latter are indicated by that the zona pellucida remains intact if an unfertilized egg is placed in the uterus under the same conditions.[7] A substance probably involved is plasmin. Plasminogen, the plasmin precursor, is found in the uterine cavity, and blastocyst factors contribute to its conversion to active plasmin. This hypothesis is supported by lytic effects in vitro by plasmin.[7] Furthermore, plasmin inhibitors also inhibit the entire zona hatching in rat experiments.[7]
Apposition
The very first, albeit loose, connection between the blastocyst and the endometrium is called the apposition.[7]
Location
On the endometrium, the apposition is usually made where there is a small crypt in it, perhaps because it increases the area of contact with the rather spherical blastocyst.
On the blastocyst, on the other hand, it occurs at a location where there has been enough lysis of the zona pellucida to have created a rupture to enable direct contact between the underlying trophoblast and the decidua of the endometrium.[7] However, ultimately, the inner cell mass, inside the trophoblast layer, is aligned closest to the decidua. Nevertheless, the apposition on the blastocyst is not dependent on if it is on the same side of the blastocyst as the inner cell mass. Rather, the inner cell mass rotates inside the trophoblast to align to the apposition.[7] In short, the entire surface of the blastocyst has a potential to form the apposition to the decidua.
Molecular Mechanism
The identity of the molecules on the trophoblast and the endometrial epithelia that mediate the initial interaction between the two remain unidentified. However, a number of research groups have proposed that MUC1, a member of the Mucin family of glycosylated proteins, is involved.[8] MUC1 is a transmembrane glycoprotein expressed at the apical surface of endometrial epithelial cells during the window of implantation in humans and has been shown to be differentially expressed between fertile and infertile subjects during this time.[8] MUC1 displays carbohydrate moieties on its extracellular domain that are ligands of L-selectin, a protein expressed on the surface of trophoblast cells.[9] An in vitro model of implantation developed by Genbacev et al., gave evidence to support the hypothesis that L-selectin mediates apposition of the blastocyst to the uterine epithelium by interacting with its ligands.[10]
Adhesion
Adhesion is a much stronger attachment to the endometrium than the loose apposition.
The trophoblasts adhere by penetrating the endometrium, with protrusions of trophoblast cells.
Communication
There is massive communication between the blastocyst and the endometrium at this stage. The blastocyst signals to the endometrium to adapt further to its presence, e.g. by changes in the cytoskeleton of decidual cells. This, in turn, dislodges the decidual cells from their connection to the underlying basal lamina, which enables the blastocyst to perform the succeeding invasion.[7]
This communication is conveyed by receptor-ligand-interactions, both integrin-matrix and proteoglycan ones.
proteoglycan receptors
Another ligand-receptor system involved in adhesion is proteoglycan receptors, found on the surface of the decidua of the uterus. Their counterparts, the proteoglycans, are found around the trophoblast cells of the blastocyst. This ligand-receptor system also is present just at the implantation window.[7]
Invasion
Invasion is an even further establishment of the blastocyst in the endometrium.
Syncytiotrophoblasts
The protrusions of trophoblast cells that adhere into the endometrium continue to proliferate and penetrate into the endometrium. As these trophoblast cells penetrate, they differentiate to become a new type of cells, syncytiotrophoblast. The prefix syn- refers to the transformation that occurs as the boundaries between these cells disappear to form a single mass of many cell nuclei (a syncytium). The rest of the trophoblasts, surrounding the inner cell mass, are hereafter called cytotrophoblasts.
Invasion continues with the syncytiotrophoblasts reaching the basal membrane beneath the decidual cells, penetrating it and further invading into the uterine stroma. Finally, the whole embryo is embedded in the endometrium. Eventually, the syncytiotrophoblasts come into contact with maternal blood and form chorionic villi. This is the initiation of forming the placenta.
Secretions
The blastocyst secretes factors for a multitude of purposes during invasion. It secretes several autocrine factors, targeting itself and stimulating it to further invade the endometrium.[7] Furthermore, secretions loosen decidual cells from each other, prevent the embryo from being rejected by the mother, trigger the final decidualization and prevent menstruation.
Autocrine
Human chorionic gonadotropin is an autocrine growth factor for the blastocyst.[7] Insulin-like growth factor 2,[7] on the other hand, stimulates the invasiveness of it.
Dislodging
The syncytiotrophoblasts dislodges decidual cells in their way, both by degradation of cell adhesion molecules linking the decidual cells together as well as degradation of the extracellular matrix between them.
Cell adhesion molecules are degraded by syncytiotrophoblast secretion of Tumor necrosis factor-alpha. This inhibits the expression of cadherins and beta-catenin.[7] Cadherins are cell adhesion molecules, and beta-catenin helps to anchor them to the cell membrane. Inhibited expression of these molecules thus loosens the connection between decidual cells, permitting the syncytotrophoblasts and the whole embryo with them to invade into the endometrium.
The extracellular matrix is degraded by serine endopeptidases and metalloproteinases. Examples of such metalloproteinases are collagenases, gelatinases and stromelysins.[7] These collagenases digest Type-I collagen, Type-II collagen, Type-III collagen, Type-VII collagen and Type-X collagen.[7] The gelatinases exist in two forms; one digesting Type-IV collagen and one digesting gelatin.[7]
Immunosuppressive
The embryo differs from the cells of the mother, and would be rejected as a parasite by the immune system of the mother if it didn't secrete immunosuppressive agents. Such agents are Platelet-activating factor, human chorionic gonadotropin, early pregnancy factor, immunosuppressive factor, Prostaglandin E2, Interleukin 1-alpha, Interleukin 6, interferon-alpha, leukemia inhibitory factor and Colony-Stimulating Factor.
Decidualization
Factors from the blastocyst also trigger the final formation of decidual cells into their proper form. In contrast, some decidual cells in the proximity of the blastocyst degenerate, providing nutrients for it.[7]
Prevention of menstruation
Human chorionic gonadotropin (hCG) not only acts as an immunosuppressive,[7] but also "notifies" the mother's body that she is pregnant, preventing menstruation by sustaining the function of the corpus luteum.
Other factors
Other factors secreted by the blastocyst are;
- Embryo-derived histamine-releasing factor
- Tissue plasminogen activator as well as its inhibitors
- Estradiol
- β1-integrins
- Fibroblast growth factor
- CYTL1[11]
- Transforming growth factor alpha
- inhibin
Failure
Implantation failure is considered to be caused by inadequate uterine receptivity in two-thirds of cases, and by problems with the embryo itself in the other third.[12]
Inadequate uterine receptivity may be caused by abnormal cytokine and hormonal signaling as well as epigenetic alterations.[13] Recurrent implantation failure is a cause of female infertility. Therefore, pregnancy rates can be improved by optimizing endometrial receptivity for implantation.[13] Evaluation of implantation markers may help to predict pregnancy outcome and detect occult implantation deficiency.[13]
Luteal support is the administration of medication, generally progestins, for the purpose of increasing the success rate of implantation and early embryogenesis, thereby complementing the function of the corpus luteum.
In women with more than 3 implantation failures in assisted reproduction, a review of several small randomized controlled studies estimated that the use of adjunct low molecular weight heparin (LMWH) improves live birth rate by approximately 80%.[14]
See also
References
- ↑ http://www.stfm.org/fmhub/fm2004/November/Walter690.pdf
- ↑ Wilcox AJ, Baird DD, Weinberg CR (1999). "Time of implantation of the Conceptus and loss of pregnancy". New England Journal of Medicine. 340 (23): 1796–1799. doi:10.1056/NEJM199906103402304. PMID 10362823.
- ↑ Xiao, Y.; Sun, X.; Yang, X.; Zhang, J.; Xue, Q.; Cai, B.; Zhou, Y. (2010). "Leukemia inhibitory factor is dysregulated in the endometrium and uterine flushing fluid of patients with adenomyosis during implantation window". Fertility and Sterility. 94 (1): 85–89. doi:10.1016/j.fertnstert.2009.03.012. PMID 19361790.
- ↑ Aboubakr M. Elnashar, Gamal I. Aboul-Enein. Endometrial receptivity. Middle East Fertility Society Journal, Vol. 9, No. 1, 2004, pp. 10-24
- ↑ 6.2 Implantation stages from embryology.ch at by the universities of Fribourg, Lausanne and Bern (Switzerland). Retrieved May, 2012
- ↑ "Implantation stages". Human Embryology. Online course in embryology for medicine students developed by the universities of Fribourg, Lausanne and Bern (Switzerland) with the support of the Swiss Virtual Campus. Retrieved 6 December 2011.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 Boron, Walter; Emile Boulpaep (2004). Medical Physiology: A Cellular And Molecular Approaoch. Oxford: Elsevier. ISBN 1-4160-2328-3. OCLC 61527528.
- 1 2 Margarit, L.; Taylor, A.; Roberts, M. H.; Hopkins, L.; Davies, C.; Brenton, A. G.; Conlan, R. S.; Bunkheila, A.; Joels, L.; White, J. O.; Gonzalez, D. (2010). "MUC1 as a Discriminator between Endometrium from Fertile and Infertile Patients with PCOS and Endometriosis". The Journal of Clinical Endocrinology & Metabolism. 95 (12): 5320–5329. doi:10.1210/jc.2010-0603. ISSN 0021-972X.
- ↑ Carson, D. D.; et al., (2006). "MUC1 is a scaffold for selectin ligands in the human uterus.". Front. Biosci. 11 (1): 2903. doi:10.2741/2018.
- ↑ Genbacev, O. D.; et al., (2003). "Trophoblast L-selectin-mediated adhesion at the maternal-fetal interface.". Science. 299: 405. doi:10.1126/science.1079546.
- ↑ Ai, Z., W. Jing, and L. Fang. "Cytokine-Like Protein 1 (Cytl1): A Potential Molecular Mediator in Embryo Implantation." PloS one 11.1 (2015): e0147424-e0147424.
- ↑ Melford, S. E.; Taylor, A. H.; Konje, J. C. (2013). "Of mice and (wo)men: factors influencing successful implantation including endocannabinoids". Human Reproduction Update. 20 (3): 415–428. doi:10.1093/humupd/dmt060. ISSN 1355-4786.
- 1 2 3 Cakmak, H.; Taylor, H. S. (2010). "Implantation failure: Molecular mechanisms and clinical treatment". Human Reproduction Update. 17 (2): 242–253. doi:10.1093/humupd/dmq037. PMC 3039220
 . PMID 20729534.
. PMID 20729534. - ↑ Potdar, N.; Gelbaya, T. A.; Konje, J. C.; Nardo, L. G. (2013). "Adjunct low-molecular-weight heparin to improve live birth rate after recurrent implantation failure: A systematic review and meta-analysis". Human Reproduction Update. 19 (6): 674–684. doi:10.1093/humupd/dmt032. PMID 23912476.