Seahorse
| Seahorses Temporal range: 23–0 Ma
| |
|---|---|
 | |
| Hippocampus sp. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Actinopterygii |
| Order: | Syngnathiformes |
| Family: | Syngnathidae |
| Subfamily: | Hippocampinae |
| Genus: | Hippocampus Rafinesque, 1810[1] |
| Species | |
|
see Species. | |
Seahorse is the name given to 54 species of small marine fishes in the genus Hippocampus. "Hippocampus" comes from the Ancient Greek word hippos meaning "horse" and kampos meaning "sea monster".[2] The word "seahorse" can also be written as two separate words (sea horse), or hyphenated (sea-horse).[3] Having a head and neck suggestive of a horse, seahorses also feature segmented bony armour, an upright posture and a curled prehensile tail.[4]
Habitat
Seahorses are mainly found in shallow tropical and temperate waters throughout the world, from about 45°S to 45°N [5] and live in sheltered areas such as seagrass beds, estuaries, coral reefs, or mangroves. Four species are found in Pacific waters from North America to South America. In the Atlantic, H. erectus ranges from Nova Scotia to Uruguay. H. zosterae, known as the dwarf seahorse, is found in the Bahamas.
Colonies have been found in European waters such as the Thames Estuary.[6]
Three species live in the Mediterranean Sea: H. guttulatus (the long-snouted seahorse), H. hippocampus (the short-snouted seahorse), and H. fuscus (the sea pony). These species form territories; males stay within 1 m2 (11 sq ft) of habitat, while females range about one hundred times that.
Description
.jpg)
Seahorses range in size from 1.5 to 35.5 cm (0.6 to 14.0 in).[7] They are named for their equine appearance with bent necks and long snouted heads followed by their distinctive trunk and tail. Although they are bony fish, they do not have scales, but rather thin skin stretched over a series of bony plates, which are arranged in rings throughout their bodies. Each species has a distinct number of rings.[8] Seahorses swim upright, another characteristic not shared by their close pipefish relatives, which swim horizontally. Razorfish are the only other fish that swim vertically. They swim upright propelling themselves by using the dorsal fin. The pectoral fins located on either side of the head are used for maneuvering. They lack the caudal fin typical of fishes. Their prehensile tail can only be unlocked in the most extreme conditions. They are adept at camouflage with the ability to grow and reabsorb spiny appendages depending on their habitat.[9]
Unusual among fish, a seahorse has a flexible, well-defined neck. It also sports a crown-like spine or horn on its head, termed a "coronet," which is distinct for each species.
Seahorses swim very poorly, rapidly fluttering a dorsal fin and using pectoral fins (located behind their eyes) to steer. The slowest-moving fish in the world is H. zosterae (the dwarf seahorse), with a top speed of about 5 ft (1.5 m) per hour.[10] Since they are poor swimmers, they are most likely to be found resting with their prehensile tails wound around a stationary object. They have long snouts, which they use to suck up food, and their eyes can move independently of each other like those of a chameleon.[11]
Evolution and fossil record
Anatomical evidence, supported by molecular, physical, and genetic evidence, demonstrates seahorses are highly modified pipefish. The fossil record of seahorses, however, is very sparse. The best known and best studied fossils are specimens of H. guttulatus (though literature more commonly refers to them under the synonym of H. ramulosus), from the Marecchia River Formation of Rimini Province, Italy, dating back to the Lower Pliocene, about 3 million years ago. The earliest known seahorse fossils are of two pipefish-like species, H. sarmaticus and H. slovenicus from the coprolitic horizon of Tunjice Hills, a middle Miocene lagerstätte in Slovenia dating back about 13 million years.[12] Molecular dating finds that pipefish and seahorses diverged during the Late Oligocene. This has led to speculation that seahorses evolved in response to large areas of shallow water, newly created as the result of tectonic events. The shallow water would have allowed the expansion of seagrass habitats that selected for the camouflage offered by the seahorses’ upright posture.[13] These tectonic changes occurred in the western Pacific Ocean, pointing to an origin there, with molecular data suggesting two later, separate invasions of the Atlantic Ocean.[14]
Reproduction
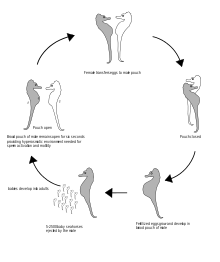
The male seahorse is equipped with a pouch on the ventral, or front-facing, side of the tail. When mating, the female seahorse deposits up to 1,500 eggs in the male's pouch. The male carries the eggs for 9 to 45 days until the seahorses emerge fully developed, but very small. Once the young are released into the water, the male's role is done and he offers no further care and often mates again within hours or days during the breeding season.[15]
Courtship
Before breeding, seahorses may court for several days. Scientists believe the courtship behavior synchronizes the animals' movements and reproductive states so the male can receive the eggs when the female is ready to deposit them. During this time, they may change color, swim side by side holding tails or grip the same strand of sea grass with their tails, and wheel around in unison in what is known as a "predawn dance". They eventually engage in a "true courtship dance" lasting about 8 hours, during which the male pumps water through the egg pouch on his trunk which expands and opens to display its emptiness. When the female’s eggs reach maturity, she and her mate let go of any anchors and drift upward snout-to-snout, out of the seagrass, often spiraling as they rise. They interact for about 6 minutes, reminiscent of courtship. The female then swims away until the next morning, and the male returns to sucking up food through his snout.[16] The female inserts her ovipositor into the male’s brood pouch and deposits dozens to thousands of eggs. As the female releases her eggs, her body slims while his swells. Both animals then sink back into the seagrass and she swims away.
Fertilization
During fertilization in Hippocampus kuda the brood pouch was found to be open only for six seconds while egg deposition occurred. During this time seawater entered the pouch where the spermatozoa and eggs meet in a seawater milieu. This hyperosmotic environment facilitates sperm activation and motility. The fertilization is therefore regarded as being physiologically ‘external’ within a physically ‘internal’ environment after the closure of the pouch.[17] It is believed that this protected form of fertilization reduces sperm competition among males. Within the Syngnathidae (pipefishes and seahorses) protected fertilization has not been documented in the pipefishes but the lack of any distinct differences in the relation of testes size to body size suggests that pipefishes may also have evolved mechanisms for more efficient fertilization with reduced sperm competition.[18]
Gestation

The fertilized eggs are then embedded in the pouch wall and become surrounded by a spongy tissue.[19] The male supplies the eggs with prolactin, the same hormone responsible for milk production in pregnant mammals. The pouch provides oxygen, as well as a controlled environment incubator. Though the egg yolk contribute nourishment to the developing embryo, the male sea horses contribute additional nutrients such as energy- rich lipids and also calcium to allow them to build their skeletal system, by secreting them into the brood pouch that are absorbed by the embryos. Further they also offer immunological protection, osmoregulation, gas exchange and waste transport [20]
The eggs then hatch in the pouch, where the salinity of the water is regulated; this prepares the newborns for life in the sea.[16][21][22] Throughout gestation, which in most species requires two to four weeks, his mate visits him daily for “morning greetings”.
Birth
The number of young released by the male seahorse averages 100–1000 for most species, but may be as low as 5 for the smaller species, or as high as 2,500. When the fry are ready to be born, the male expels them with muscular contractions. He typically gives birth at night and is ready for the next batch of eggs by morning when his mate returns. Like almost all other fish species, seahorses do not nurture their young after birth. Infants are susceptible to predators or ocean currents which wash them away from feeding grounds or into temperatures too extreme for their delicate bodies. Less than 0.5% of infants survive to adulthood, explaining why litters are so large. These survival rates are actually fairly high compared to other fish, because of their protected gestation, making the process worth the great cost to the father. The eggs of most other fish are abandoned immediately after fertilization.[22]
Questions surrounding reproductive roles
Reproduction is energetically costly to the male. This brings into question why the sexual role reversal even takes place. In an environment where one partner incurs more energy costs than the other, Bateman's principle suggests that the lesser contributor takes the role of the aggressor. Male seahorses are more aggressive and sometimes “fight” for female attention. According to Amanda Vincent of Project Seahorse, only males tail-wrestle and snap their heads at each other. This discovery prompted further study of energy costs. To estimate the female’s direct contribution, researcher Heather D. Masonjones, associate professor of biology at the University of Tampa, chemically analyzed the energy stored in each egg. To measure the burden on the male, Masonjones measured its oxygen consumption. By the end of incubation, the male consumed almost 33% more oxygen than before mating. The study concluded that the female's energy expenditure while generating eggs is twice that of males during incubation[16] confirming the standard hypothesis.
Why the male seahorse (and other members of the Syngnathidae) carries the offspring through gestation is unknown, though some researchers [23] believe it allows for shorter birthing intervals, in turn resulting in more offspring. Given an unlimited number of ready and willing partners, males have the potential to produce 17% more offspring than females in a breeding season. Also, females have “time-outs” from the reproductive cycle 1.2 times longer than those of males. This seems to be based on mate choice, rather than physiology. When the female’s eggs are ready, she must lay them in a few hours or eject them into the water column. Making eggs is a huge cost to her physically, since they amount to about a third of her body weight. To protect against losing a clutch, the female demands a long courtship. The daily greetings help to cement the bond between the pair.[24]
Monogamy
One common misconception about seahorses is that they mate for life. Many species of seahorses form pair bonds that last through at least the breeding season. Some species show a higher level of mate fidelity than others.[25][26] However, many species readily switch mates when the opportunity arises. H. abdominalis and H. breviceps have been shown to breed in groups, showing no continuous mate preference. Many more species' mating habits have not been studied, so it is unknown how many species are actually monogamous, or how long those bonds actually last.[27]
Although monogamy within fish is not common, it does appear to exist for some. In this case, the mate-guarding hypothesis may be an explanation. This hypothesis states, “males remain with a single female because of ecological factors that make male parental care and protection of offspring especially advantageous.”[28] Because the rates of survival for newborn seahorses are so low, incubation is essential. Though not proven, males could have taken on this role because of the lengthy period the females require to produce their eggs. If males incubate while females prepare the next clutch (amounting to a third of body weight), they can reduce the interval between clutches.
Feeding habits

Seahorses feed on small crustaceans floating in the water or crawling on the bottom. With excellent camouflage and patience, seahorses ambush prey that floats within striking range. Mysid shrimp and other small crustaceans are favorites, but some seahorses have been observed eating other kinds of invertebrates and even larval fish. In a study of seahorses, the distinctive head morphology was found to give it a hydrodynamic advantage that creates minimal interference while approaching an evasive prey. Therefore, the seahorse has the ability to come within a very close range of copepods, on which they prey.[29][30] After successfully closing in on the prey without alerting it, the sea horse gives an upward thrust rapidly rotates the head aided by large tendons that store and release elastic energy, to bring its long snout close to the prey.This step is crucial for prey capture as suction only works with the mouth at a close range. This two phase prey capture mechanism is termed pivot-feeding.[30][31]
While feeding, they produce a distinctive click each time a food item is ingested. The same clicks are heard with social interactions.
Threats of extinction
Because data is lacking on the sizes of the various seahorse populations, as well as other issues including how many seahorses are dying each year, how many are being born, and the number used for souvenirs, there is insufficient information to assess their risk of extinction, and the risk of losing more seahorses remains a concern. Some species, such as the Paradoxical Seahorse, H. paradoxus,[32] may already be extinct. Coral reefs and seagrass beds are deteriorating, reducing viable habitats for seahorses.[33]
Aquaria

While many aquarium hobbyists keep them as pets, seahorses collected from the wild tend to fare poorly in home aquaria. Many eat only live foods such as brine shrimp and are prone to stress, which damages their immune systems and makes them susceptible to disease.

In recent years, however, captive breeding has become more popular. Such seahorses survive better in captivity, and are less likely to carry diseases. They eat frozen mysidacea (crustaceans) that are readily available from aquarium stores,[34] and do not experience the stress of moving out of the wild. Although captive-bred seahorses are more expensive, they take no toll on wild populations.
Seahorses should be kept in an aquarium with low flow and placid tank mates. They are slow feeders, so fast, aggressive feeders will leave them without food.[34] Seahorses can coexist with many species of shrimp and other bottom-feeding creatures. Gobies also make good tank-mates. Keepers are generally advised to avoid eels, tangs, triggerfish, squid, octopus, and sea anemones.[35]
Water quality is very important for the survival of seahorses in an aquarium. They are delicate species which should not be added to a new tank. The water parameters are recommended to be as follows although these fish may acclimatise to different water over time: Temperature: 23–28 °C (73–82 °F) pH: 8.1–8.4 Ammonia: 0 mg/l (0 ppm) (0.01 mg/l (0.01 ppm) may be tolerated for short periods) Nitrite: 0 mg/l (0 ppm) (0.125 mg/l (0.125 ppm) may be tolerated for short periods) S.G.: 1.021–1.024 at 23–24 °C (73–75 °F) A water-quality problem will affect fish behaviour and can be shown by clamped fins, reduced feeding, erratic swimming, and gasping at the surface.[36] Seahorses swim up and down, as well as using the length of the aquarium. Therefore, the tanks should ideally be twice as deep as the length of the adult seahorse.
Animals sold as "freshwater seahorses" are usually the closely related pipefish, of which a few species live in the lower reaches of rivers. The supposed true "freshwater seahorse" called H. aimei is not a valid species, but a synonym sometimes used for Barbour's and hedgehog seahorses. The latter, which is often confused with the former, can be found in estuarine environments, but is not actually a freshwater fish.[37]
Use in Chinese medicine

Seahorse populations are thought to be endangered as a result of overfishing and habitat destruction. Despite a lack of scientific studies or clinical trials,[38][39] the consumption of seahorses is widespread in traditional Chinese medicine, primarily in connection with impotence, wheezing, nocturnal enuresis, and pain, as well as labor induction.[40] Up to 20 million seahorses may be caught each year to be sold for such uses.[41] Preferred species of seahorses include H. kellogii, H. histrix, H. kuda, H. trimaculatus, and H. mohnikei.[40] Seahorses are also consumed by the Indonesians, the central Filipinos, and many other ethnic groups.
Import and export of seahorses has been controlled under CITES since 15 May 2004. However, Indonesia, Japan, Norway, and South Korea have chosen to opt out of the trade rules set by CITES.
The problem may be exacerbated by the growth of pills and capsules as the preferred method of ingesting seahorses. Pills are cheaper and more available than traditional, individually tailored prescriptions of whole seahorses, but the contents are harder to track. Seahorses once had to be of a certain size and quality before they were accepted by TCM practitioners and consumers. Declining availability of the preferred large, pale, and smooth seahorses has been offset by the shift towards prepackaged preparations, which makes it possible for TCM merchants to sell previously unused, or otherwise undesirable juvenile, spiny, and dark-coloured animals. Today, almost a third of the seahorses sold in China are packaged, adding to the pressure on the species.[42]
Dried seahorse retails from US$600 to $3000 per kilogram, with larger, paler, and smoother animals commanding the highest prices. In terms of value based on weight, seahorses retail for more than the price of silver and almost that of gold in Asia.[43]
Species
As of 2012 the 54 recognized species in this genus are:[44]
.jpg)


- Hippocampus abdominalis Lesson, 1827 (big-belly seahorse)
- Hippocampus alatus Kuiter, 2001 (winged seahorse)
- Hippocampus algiricus Kaup, 1856 (West African seahorse)
- Hippocampus angustus Günther, 1870 (narrow-bellied seahorse)
- Hippocampus barbouri D. S. Jordan & R. E. Richardson, 1908 (Barbour's seahorse)
- Hippocampus bargibanti Whitley, 1970 (pygmy seahorse)
- Hippocampus biocellatus Kuiter, 2001 (false-eyed seahorse)
- Hippocampus borboniensis A. H. A. Duméril, 1870 (Réunion seahorse)
- Hippocampus breviceps W. K. H. Peters, 1869 (short-headed seahorse)
- Hippocampus camelopardalis Bianconi, 1854 (giraffe seahorse)
- Hippocampus capensis Boulenger, 1900 (Knysna seahorse)
- Hippocampus colemani Kuiter, 2003
- Hippocampus comes Cantor, 1850 (tiger-tail seahorse)
- Hippocampus coronatus Temminck & Schlegel, 1850 (crowned seahorse)
- Hippocampus curvicuspis R. Fricke, 2004 (New Caledonian thorny seahorse)
- Hippocampus debelius M. F. Gomon & Kuiter, 2009 (softcoral seahorse)
- Hippocampus denise Lourie & J. E. Randall, 2003 (Denise's pygmy seahorse)
- Hippocampus erectus Perry, 1810 (lined seahorse)
- Hippocampus fisheri D. S. Jordan & Evermann, 1903 (Fisher's seahorse)
- Hippocampus fuscus Rüppell, 1838 (sea pony)
- Hippocampus grandiceps Kuiter, 2001 (big-head seahorse)
- Hippocampus guttulatus G. Cuvier, 1829 (long-snouted seahorse)
- Hippocampus hendriki Kuiter, 2001 (eastern spiny seahorse)
- Hippocampus hippocampus (Linnaeus, 1758) (short-snouted seahorse)
- Hippocampus histrix Kaup, 1856 (spiny seahorse)
- Hippocampus ingens Girard, 1858 (Pacific seahorse)
- Hippocampus jayakari Boulenger, 1900 (Jayakar's seahorse)
- Hippocampus jugumus Kuiter, 2001 (collared seahorse)
- Hippocampus kelloggi D. S. Jordan & Snyder, 1901 (great seahorse)
- Hippocampus kuda Bleeker, 1852 (spotted seahorse)
- Hippocampus lichtensteinii Kaup, 1856 (Lichtenstein's seahorse)
- Hippocampus minotaur M. F. Gomon, 1997 (bullneck seahorse)
- Hippocampus mohnikei Bleeker, 1854 (Japanese seahorse)
- Hippocampus montebelloensis Kuiter, 2001 (Monte Bello seahorse)
- Hippocampus multispinus Kuiter, 2001 (northern spiny seahorse)
- Hippocampus paradoxus Foster & M. F. Gomon, 2010 (paradoxical seahorse)
- Hippocampus patagonicus Piacentino & Luzzatto, 2004
- Hippocampus pontohi Lourie & Kuiter, 2008
- Hippocampus procerus Kuiter, 2001 (high-crown seahorse)
- Hippocampus pusillus R. Fricke, 2004 (pygmy thorny seahorse)
- Hippocampus queenslandicus Horne, 2001 (Queensland seahorse)
- Hippocampus reidi Ginsburg, 1933 (longsnout seahorse)
- Hippocampus satomiae Lourie & Kuiter, 2008 (Satomi's pygmy seahorse)
- Hippocampus semispinosus Kuiter, 2001 (half-spined seahorse)
- Hippocampus severnsi Lourie & Kuiter, 2008
- Hippocampus sindonis D. S. Jordan & Snyder, 1901 (Dhiho's seahorse)
- Hippocampus spinosissimus M. C. W. Weber, 1913 (hedgehog seahorse)
- Hippocampus subelongatus Castelnau, 1873 (West Australian seahorse)
- Hippocampus trimaculatus Leach, 1814 (longnose seahorse)
- Hippocampus tyro J. E. Randall & Lourie, 2009
- Hippocampus waleananus M. F. Gomon & Kuiter, 2009 (Walea pygmy seahorse)
- Hippocampus whitei Bleeker, 1855 (White's seahorse)
- Hippocampus zebra Whitley, 1964 (zebra seahorse)
- Hippocampus zosterae D. S. Jordan & C. H. Gilbert, 1882 (dwarf seahorse)

Pygmy seahorses
Pygmy seahorses are less than 15 millimeters (0.59 in) tall and 17 millimeters (0.67 in) wide members of the genus. Previously the term was applied exclusively to the species H. bargibanti but since 1997, discoveries have made this term obsolete. The species H. minotaur, H. denise, H. colemani, H. pontohi, H. severnsi, H. satomiae and H. waleananus have been described. Other species that are believed to be unclassified have also been reported in books, dive magazines and on the Internet. They can be distinguished from other species of seahorse by their 12 trunk rings, low number of tail rings (26–29), the location in which young are brooded in the trunk region of males and their extremely small size.[45] Molecular analysis (of ribosomal RNA) of 32 Hippocampus species found that H. bargibanti belongs in a separate clade from other members of the genus and therefore that the species diverged from the other species in the "ancient" past.[46]
Most pygmy seahorses are well camouflaged and live in close association with other organisms including colonial hydrozoans (Lytocarpus and Antennellopsis), coralline algae (Halimeda) sea fans (Muricella, Annella, Acanthogorgia). This combined with their small size accounts for why most species have only been noticed and classified since 2001.[45][47]
References
- ↑ Hippocampus Rafinesque, 1810, WoRMS
- ↑ Duvernoy, HM (2005). "Introduction". The Human Hippocampus (3rd ed.). Berlin: Springer-Verlag. p. 1. ISBN 3-540-23191-9.
- ↑ Shorter Oxford English Dictionary. Oxford, UK: Oxford University Press. 2007. ISBN 0199206872.
- ↑ "sea horse or seahorse". dictionary.com. Retrieved 2016-06-19.
- ↑ "Home". Project Seahorse. Retrieved 2015-11-15.
- ↑ "Rare seahorses breeding in Thames". BBC News. 7 April 2008. Retrieved 11 November 2009.
- ↑ "Seahorses, Seahorse Pictures, Seahorse Facts". National Geographic. Retrieved 17 May 2012.
- ↑ "Observatoire Océanologique de Banyuls sur mer". www.obs-banyuls.fr. Retrieved 2015-11-16.
- ↑ Garrick-Maidment, N.; Trewhella, S.; Hatcher, J.; Collins, K.j.; Mallinson, J.j. (2010-01-01). "Seahorse Tagging Project, Studland Bay, Dorset, UK". Marine Biodiversity Records. 3. doi:10.1017/S175526721000062X. ISSN 1755-2672.
- ↑ Guinness Book of World Records (2009)
- ↑ Lourie, Sara. Seahorses: A Life-size Guide to Every Species. Ivy Press. ISBN 9781782403210.
- ↑ Žalohar J.; Hitij T.; Križnar M. (2009). "Two new species of seahorses (Syngnathidae, Hippocampus) from the Middle Miocene (Sarmatian) Coprolitic Horizon in Tunjice Hills, Slovenia: The oldest fossil record of seahorses". Annales de Paléontologie. 95 (2): 71–96. doi:10.1016/j.annpal.2009.03.002.
- ↑ Teske PR; Beheregaray LB (2009). "Evolution of seahorses' upright posture was linked to Oligocene expansion of seagrass habitats". Biol Lett. 5 (4): 521–3. doi:10.1098/rsbl.2009.0152. PMC 2781918
 . PMID 19451164.
. PMID 19451164. - ↑ Teske PR; Cherry MI; Matthee CA (2004). "The evolutionary history of seahorses (Syngnathidae: Hippocampus): molecular data suggest a West Pacific origin and two invasions of the Atlantic Ocean". Mol Phylogenet Evol. 30 (2): 273–86. doi:10.1016/S1055-7903(03)00214-8. PMID 14715220.
- ↑ Foster S.J; Vincent C.J. (2004). "Life history and ecology of seahorses: implications for conservation and management". Journal of Fish Biology. 65: 1–61. doi:10.1111/j.1095-8649.2004.00429.x.
- 1 2 3 Milius, S. (2000). "Pregnant: And Still Macho" (PDF). Science News. 157 (11): 168. doi:10.2307/4012130. JSTOR 4012130.
- ↑ Look, Katrien J. W. Van; Dzyuba, Borys; Cliffe, Alex; Koldewey, Heather J.; Holt, William V. (2007-02-01). "Dimorphic sperm and the unlikely route to fertilisation in the yellow seahorse". Journal of Experimental Biology. 210 (3): 432–437. doi:10.1242/jeb.02673. ISSN 0022-0949. PMID 17234612.
- ↑ Kvarnemo, Charlotta; Simmons, Leigh W. (2004). "Testes investment and spawning mode in pipefishes and seahorses (Syngnathidae)". Biological Journal of the Linnean Society. 83: 369–376. doi:10.1111/j.1095-8312.2004.00395.x.
- ↑ "The biology of seahorses: Reproduction". The Seahorse Project. Archived from the original on 3 March 2009. Retrieved 8 May 2007.
- ↑ Whittington, Camilla M.; Griffith, Oliver W.; Qi, Weihong; Thompson, Michael B.; Wilson, Anthony B. (2015-09-01). "Seahorse Brood Pouch Transcriptome Reveals Common Genes Associated with Vertebrate Pregnancy". Molecular Biology and Evolution. 32: msv177. doi:10.1093/molbev/msv177. ISSN 0737-4038. PMID 26330546.
- ↑ Masonjones, H. D.; Lewis, S. M. (2000). "Differences in potential reproductive rates of male and female seahorses related to courtship roles". Animal Behaviour. 59 (1): 11–20. doi:10.1006/anbe.1999.1269. PMID 10640362.
- 1 2 Danielson, Stentor (14 June 2002). "Seahorse Fathers Take Reins in Childbirth". National Geographic News.
- ↑ Vincent, Amanda C. J. (1994). "Operational Sex Ratios in Seahorses". Behaviour. 128 (1/2): 153–167. doi:10.1163/156853994X00091. JSTOR 4535169.
- ↑ "Why Do Male Seahorses Get Pregnant?". Petseahorse.com.
- ↑ Kvarnemo C; Moore G.I; Jones A.G; Nelson W.S; Avise J.C. (2000). "Monogamous pair bonds and mate switching in the Western Australian seahorse Hippocampus subelongatus". J. Evol. Biol. 13 (6): 882–8. doi:10.1046/j.1420-9101.2000.00228.x.
- ↑ Vincent C.J.; Sadler L.M. (1995). "Faithful pair bonds in wild seahorses, Hippocampus whitei" (PDF). Anim. Behav. 50 (6): 1557–1569. doi:10.1016/0003-3472(95)80011-5. Archived from the original (PDF) on 23 July 2011.
- ↑ Weiss, Tami (10 April 2010). "What's Love Got to Do With It? The Truth About Seahorse Monogamy". fusedjaw.com.
- ↑ Alcock, John (2005). Animal Behavior (8th ed.). Massachusetts: Sinauer. pp. 370–1. ISBN 0878930051.
- 1 2 Langley, Liz (26 November 2013). "Why Does the Seahorse Have Its Odd Head? Mystery Solved – News Watch". Newswatch.nationalgeographic.com.
- 1 2 Gemmell, B. J.; Sheng, J.; Buskey, E. J. (2013). "Morphology of seahorse head hydrodynamically aids in capture of evasive prey". Nature Communications. 4. doi:10.1038/ncomms3840.
- ↑ Wassenbergh, Sam Van; Strother, James A.; Flammang, Brooke E.; Ferry-Graham, Lara A.; Aerts, Peter (2008-03-06). "Extremely fast prey capture in pipefish is powered by elastic recoil". Journal of the Royal Society Interface. 5 (20): 285–296. doi:10.1098/rsif.2007.1124. ISSN 1742-5689. PMC 2607401. PMID 17626004.
- ↑ "New species of seahorse found... after sitting in a museum for more than a decade". Daily Mail. <!- – 09:04 EST --> 17 February 2011. Retrieved 4 April 2014. Check date values in:
|date=(help) - ↑ Lourie, Sarah A.; Foster, Sarah J.; Cooper, Ernest W.T. and Vincent, Amanda C.J. (2004) A Guide to the Identification of Seahorses. Project Seahorse Advancing Marine Conservation, ISBN 0-89164-169-6.
- 1 2 "Seahorse and Pipefish Foods | Tami Weiss". Fusedjaw.com. 25 June 2005. Retrieved 11 November 2009.
- ↑ "Seahorse Tankmates | Will Wooten". Fusedjaw.com. 25 June 2004. Retrieved 11 November 2009.
- ↑ How to care for Seahorses & Pipefish. seahorseaquariums.ie
- ↑ "Hippocampus spinosissimus". Fishbase. Retrieved 11 November 2009.
- ↑ Stephen Barrett, M.D. "Be Wary of Acupuncture, Qigong, and "Chinese Medicine"". Retrieved 11 December 2013.
- ↑ Still, J. (2003). "Use of animal products in traditional Chinese medicine: Environmental impact and health hazards". Complementary Therapies in Medicine. 11 (2): 118–22. doi:10.1016/S0965-2299(03)00055-4. PMID 12801499.
- 1 2 Bensky, D., Clavey, S., Stoger, E. (2004) Chinese Herbal Medicine: Materia Medica. Eastland Press, Inc. Seattle, 3rd ed. ISBN 0939616424. p. 815
- ↑ "Seahorse Crusader Amanda Vincent" on Nova television show
- ↑ Parry-Jones, Rob & Vincent, Amanda (3 January 1998). "Can we tame wild medicine?". New Scientist.
- ↑ "Save Our Seahorses". Save Our Seahorses. Retrieved 13 May 2014.
- ↑ Froese, Rainer, and Daniel Pauly, eds. (2012). Species of Hippocampus in FishBase. October 2012 version.
- 1 2 Lourie, Sara; Rudie Kuiter (2008). "Three new pygmy seahorse species from Indonesia (Teleostei: Syngnathidae: Hippocampus)" (PDF). Zootaxa. 1963: 54–68. ISSN 1175-5334. Retrieved 9 June 2009.
- ↑ Teske, Peter; Michael Cherry; Conrad Matthee (February 2004). "The evolutionary history of seahorses (Syngnathidae: Hippocampus): molecular data suggest a West Pacific origin and two invasions of the Atlantic Ocean". Molecular Phylogenetics and Evolution. 30 (2): 273–286. doi:10.1016/S1055-7903(03)00214-8. PMID 14715220.
- ↑ "Science in Pictures: Pygmy Seahorses." The Epoch Times, Northern California Edition (8 November 2011).
Further reading
- Amanda C.J. Vincent & Laila M. Sadler (1995). "Faithful pair bonds in wild seahorse, Hippocampus whitei". Animal Behaviour. 50 (6): 1557–69. doi:10.1016/0003-3472(95)80011-5.
- Amanda C.J. Vincent (1995). "A role for daily greetings in maintaining seahorse pair bonds". Animal Behaviour. 49: 258–260. doi:10.1016/0003-3472(95)80178-2.
- Amanda C.J. Vincent (1990). "A seahorse father makes a good mother". Natural History. 12: 34–43.
- Amanda C.J. Vincent & Rosie Woodroffe (1994). "Mothers little helpers: patterns of male care in mammals". Trends in Ecology and Evolution. 9 (8): 294–7. doi:10.1016/0169-5347(94)90033-7. PMID 21236858.
- John Sparks (1999). Battle of the Sexes: The Natural History of Sex. London: BBC Books. ISBN 0-563-37145-5.
- Sara A. Lourie, Amanda C.J. Vincent and Heather J. Hall (1999). Seahorses: An Identification Guide to the World's Species and their Conversation. London: Project Seahorse.
- Teske P. R., Hamilton H., Matthee C. A. & Barker N. P. (2007). "Signatures of seaway closures and founder dispersal in the phylogeny of a circumglobally distributed seahorse lineage". BMC Evolutionary Biology. 7: 138. doi:10.1186/1471-2148-7-138. PMC 1978501. PMID 17697373.
- Leslee Matsushige, Curator, Birch Aquarium at Scripps, UCSD and Dawn Navarro Ericson, Natural Science Illustrator "Sea Horses & their Relatives", WEEKEND NATURALIST: An Identification and Natural Science Guide (2011) ISBN 9780982835623 Manta Publications 2011 first edition.
External links
| Wikimedia Commons has media related to Seahorse. |
- Vimeo.com, Video of various seahorse species at the Monterey Bay Aquarium
- ARKive—images and movies of the pygmy seahorse (Hippocampus bargibanti)
- Seahorse at the Encyclopedia of Life
