Pseudo-nitzschia
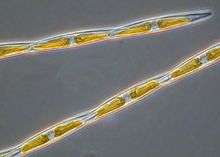
| Pseudo-nitzschia | |
|---|---|
| Scientific classification | |
| (unranked): | SAR |
| Phylum: | Heterokontophyta |
| Class: | Bacillariophyceae |
| Order: | Bacillariales |
| Family: | Bacillariaceae |
| Genus: | Pseudo-nitzschia H. Perag. in H. Perag. and Perag. |
Pseudo-nitzschia is a marine planktonic diatom genus containing some species capable of producing the neurotoxin domoic acid (DA), which is responsible for the neurological disorder known as amnesic shellfish poisoning. Currently, 46 species are known, 19 of which are known to produced DA. It was originally hypothesized that only dinoflagellates could produce harmful algal toxins, but a deadly bloom of Pseudo-nitzschia occurred in 1987 in the bays of Prince Edward Island, Canada, and led to an outbreak of ASP.[2] Over 100 people were affected by this outbreak after consuming contaminated mussels; three people died.[3] Blooms have since been characterized in coastal waters worldwide and have been linked to increasing marine nutrient concentrations.[4]
Morphology and physiology
Pseudo-nitzschia species are bilaterally symmetrical Pennate diatoms. Cell walls are made up of elongated silica frustules. The silica wall is fairly dense which leads to negative buoyancy, providing a number of advantages. The wall allows the diatoms to sink to avoid light inhibition or nutrient limitations, as well as to protect against grazing zooplankton. The silica frustules also contribute vastly to the sediment layers of the earth and to the fossil record, which makes them exceptionally useful in increasing understanding of numerous processes such as gauging the degree of climate change.[5] Before sinking to the ocean floor, every atom of silicon that enters the ocean is integrated into the cell wall of a diatom about 40 times.[5]
Silica frustules contain a central raphe, which secretes mucilage that allows the cells to move by gliding.[6] Cells are often found in overlapped, stepped colonies, and exhibit collective motility.[6] Pseudo-nitzschia species synthesize their own food through the use of light and nutrients in photosynthesis. The diatoms have a central vacuole to store nutrients for later use and a light-harvesting system to protect themselves against high-intensity light.[5]
Taxonomy
The diatom lineage may go back 180 to 250 million years ago (Mya). About 65 Mya, diatoms survived a mass extinction in which roughly 85% of all species perished.[5] Until 1994, the genus was known as Nitzschia, but was changed to Pseudo-nitzschia because of the ability to form chains of overlapping cells, as well as other minor morphological differences.[7] While the genus can be readily recognized using light microscopy, identification of distinct species can require taxonomic expertise and be extremely time-consuming. Species identification in this genus is notoriously difficult due to the presence of cryptic species. Similar species are often differentiated by very small differences in the frustule, such as shape, period, and band stria.[8]
The direct impacts of species identification on public health make this a serious concern. Toxogenic and nontoxogenic species commonly co-occur; therefore, discrimination between various Pseudo-nitzschia species is imperative to determine the potential toxicity of an algal bloom. Optical microscopy identification techniques are inadequate when a large number of samples must be routinely examined, such as is required for a monitoring program for public health. Recently, a DNA-microarray was developed for simultaneous detection of multiple harmful algal bloom species with an emphasis on Pseudo-nitzschia. The total assay is believed to have the potential to identify hundreds of species and accurately differentiate between large quantities of related species. Additionally, this technology has been shown to accurately identify toxic phytoplankton even at extremely low concentrations. The lower limit for detection of Pseudo-nitzschia is as low as 500 cells.[9]
The nomenclatural history is given in Hasle (1995)[10] and Bates (2000).[11]
Lifecycle
Ornithine-urea cycle
The physiological distribution, fixation, and recycling center for inorganic carbon and nitrogen plays a key role in the metabolic response of diatoms to prolonged nutrient deprivation. The cycle enables diatoms to respond immediately to the availability of nutrients and recover by increasing their metabolic and growth rates.[5]
Resting stages
Diatoms have the ability to enter two distinct resting stages to overcome periods of stress. A resting spore has a great capacity to survive over extended periods of nutrient deprivation. To avoid low nutrient concentrations during stratification, the resting spores can settle to the bottom where the nutrient concentration is higher. A resting cell is better able to rapidly respond when nutrients become available again. This is more often observed in freshwater and pennate diatoms like Pseudo-nitzschia.[5]
Reproduction
Among diatoms, reproduction is primarily asexual by binary fission, with each daughter cell receiving one of the parent’s cell’s two frustules. However, this asexual division results in a size reduction. To restore the cell size of a diatom population, sexual reproduction must occur. Vegetative diploid cells undergo meiosis to produce active and passive gametes. These gametes then fuse to form a zygote, which then develops into an auxospore.[12] Sexual reproduction leads to both an increase in genotypic diversity and the formation of large initial cells through formation of the auxospore. Cells need to be below a species-specific size threshold for the sexual phase to be induced. Many external cues also regulate the initiation, such as day length, irradiance, and temperature.[13]
The basic mode of the sexual phase of reproduction appears to be conserved among Pseudo-nitzschia species. Upon mixing two strains of compatible mating type and of appropriate cell size for sexualization, cells align side by side and differentiate into gametangia. One active (+) and one passive (-) gamete are then produced within each gametangium. The active gamete migrates toward the passive partner and conjugates. The zygote is then becomes an auxospore, which has no rigid frustule. Inside the auxospore, a large initial cell is produced.[12]
Sexual reproduction appears to occur exclusively in the exponential growth phase and be linked to cell density. Sexualization can only be initiated when a species-specific threshold cell concentration is met. Decreasing the distance to facilitate contact and/or perception of chemical cues between cells triggers the sexual phase, indicating that high cell density is favorable for sexual reproduction. Additionally, the onset of sexualization is linked to a significant reduction in growth of the vegetative and parental cells, suggesting that vegetative division is inhibited when the two strains of opposite mating type come in contact.[13]
Genome and transcriptome
Pseudo-nitzschia multiseries has a genome consisting of 219 megabases (Mb) and a full genome project is underway.[14]
Transcriptomes of three species, P. arenysensis, P. delicatissima, and P. multistriata, have been sequenced. The transcriptomes encode between 17,500 and 20,200 proteins. P. multistriata was found to uniquely encode nitric oxide synthase.[15]
Habitat
Pseudo-nitzschia species can be found in coastal regions worldwide. They have been documented along the Pacific coast from Canada to California, along the Atlantic Northeast coast of Canada, North Carolina, and the Gulf of Mexico.[16] Recently, they have been detected in the open ocean as well as gulfs and bays, showing a presence in many diverse environments, including off the coasts of Canada, Portugal, France, Italy, Coratia, Greece, Ireland and Australia.[17] In general, diatoms flourish in turbulent, nutrient-rich waters.[5]
Harmful bloom dynamics
Harmful algal blooms (HABs) of Pseudo-nitzschia and the like can cause diseases and death in many marine creatures, as well as the humans who consume them. The four main mechanisms through which these negative impacts can occur are:
- The increased biomass production from HABs can result in oxygen depletion.
- Toxins, like DA, can be transferred to animals higher up the food chain.
- The needle shape of diatoms can cause physical damage to skin and gills.
- The increased photosynthetic activity can lead to oxygen supersaturation.[18]
Unlike certain dinoflagellate blooms, domoic acid-producing Pseudo-nitzschia species must be present in high concentrations (greater than 100,000 cells L−1) to contaminate shellfish at a level that would cease harvesting.[19]
The largest Pseudo-nitzschia bloom ever recorded occurred in September 2004 off the northwest coast of the United States. The maximum cell densities reached 13 x 106 cells per liter, with domoic acid levels of 1.3 pg DA/cell.[20]
Sediment cores indicate a link between increasing coastal nutrient levels (eutrophication) and an increase in Pseudo-nitzschia blooms.[4]
Domoic acid
Shellfish become contaminated after feeding on toxic Pseudo-nitzschia blooms and can act as a vector to transfer domoic acid to humans upon ingestion. DA acts as a potent glutamate agonist and is responsible for amnesic shellfish poisoning in humans. Effects can be as minor as vomiting, cramps, and a headache, or as severe as permanent short-term memory loss, coma, and death.[17] So, monitoring systems and management practices for recreational and commercial fishing are important to ensure the health of marine animals and their predators.
Photosynthesis is essential for the production of domoic acid. Periods of darkness or chemical inhibition of photosynthesis has been shown to inhibit toxin production. Additionally, DA production peaks in the stationary phase of the growth cycle when cell division is slowed or absent. Production is minimal or nonexistent during the exponential phase, and ceases completely during the death phase of the growth cycle.[20]
Factors affecting production
Many factors have been linked to promotion of DA production, including sufficient light, elevated or decreased pH, and nutrition limitations. In one species, P. cuspidata, a link has been indicated between toxicity and photosynthesis photon flux density (PPFD). At a low PPFD, the exponential growth rate approximately halved and the cells were significantly more toxic.[20]
While the effect of availability of different nitrogen sources on toxicity has been studied many times, no general rule could be demonstrated for differences in growth and DA production, with the results varying greatly by species. However, toxin production increases when the nitrogen source could not sustain a high biomass, suggesting growth limitation seems to induce toxicity.[21]
The presence of zooplankton has also been shown to affect the toxicity of Pseudo-nitzschia. The presence of copepods was shown to enhance toxin production of P. seriata. This effect appears to be chemically mediated, as it could be induced without physical contact.[22]
Pseudo-nitzschia species also appear to respond dramatically to differences in trace metal concentrations, such as iron (Fe) and copper (Cu). In Fe-limited conditions, Pseudo-nitzschia increases DA production by six to 25 times as a result of stress.[2] This increase allows them to enhance Fe acquisition needed for metabolic activities, and can have devastating effects.
Known species
Known species of Pseudo-nitzschia (48) are:
- Pseudo-nitzschia abrensis
- Pseudo-nitzschia americana
- Pseudo-nitzschia antarctica
- Pseudo-nitzschia arctica
- Pseudo-nitzschia arenysensis
- Pseudo-nitzschia australis
- Pseudo-nitzschia batesiana
- Pseudo-nitzschia bipertita
- Pseudo-nitzschia brasiliana
- Pseudo-nitzschia caciantha
- Pseudo-nitzschia calliantha
- Pseudo-nitzschia circumpora
- Pseudo-nitzschia cuspidata
- Pseudo-nitzschia decipiens
- Pseudo-nitzschia delicatissima
- Pseudo-nitzschia dolorosa
- Pseudo-nitzschia fukuyoi
- Pseudo-nitzschia fraudulenta
- Pseudo-nitzschia fryxelliana
- Pseudo-nitzschia galaxiae
- Pseudo-nitzschia granii
- Pseudo-nitzschia hasleana
- Pseudo-nitzschia heimii
- Pseudo-nitzschia inflatula
- Pseudo-nitzschia kodamae
- Pseudo-nitzschia limii
- Pseudo-nitzschia linea
- Pseudo-nitzschia lineola
- Pseudo-nitzschia lundholmiae
- Pseudo-nitzschia mannii
- Pseudo-nitzschia micropora
- Pseudo-nitzschia multiseries
- Pseudo-nitzschia multistriata
- Pseudo-nitzschia obtusa
- Pseudo-nitzschia plurisecta
- Pseudo-nitzschia prolongatoides
- Pseudo-nitzschia pseudodelicatissima
- Pseudo-nitzschia pungens
- Pseudo-nitzschia pungiformis
- Pseudo-nitzschia roundii
- Pseudo-nitzschia sabit
- Pseudo-nitzschia seriata
- Pseudo-nitzschia sinica
- Pseudo-nitzschia subcurvata
- Pseudo-nitzschia subfraudulenta
- Pseudo-nitzschia subpacifica
- Pseudo-nitzschia turgidula
- Pseudo-nitzschia turgiduloides
Light and electron microscope images of Pseudo-nitzschia species are shown at the Nordic Microalgae website and scanning electron microscopy images at the Digital Microscopy Facility website.
Pseudo-nitzschia species that have been shown to produce domoic acid (23), although not all strains are toxigenic:
- Pseudo-nitzschia australis
- Pseudo-nitzschia abrensis
- Pseudo-nitzschia batesiana
- Pseudo-nitzschia brasiliana
- Pseudo-nitzschia caciantha
- Pseudo-nitzschia calliantha
- Pseudo-nitzschia cuspidata
- Pseudo-nitzschia delicatissima
- Pseudo-nitzschia fraudulenta
- Pseudo-nitzschia fukuyoi
- Pseudo-nitzschia galaxiae
- Pseudo-nitzschia granii
- Pseudo-nitzschia kodamae
- Pseudo-nitzschia lundholmiae
- Pseudo-nitzschia multiseries
- Pseudo-nitzschia multistriata
- Pseudo-nitzschia plurisecta
- Pseudo-nitzschia pungens
- Pseudo-nitzschia pseudodelicatissima
- Pseudo-nitzschia seriata
- Pseudo-nitzschia subfraudulenta
- Pseudo-nitzschia subpacifica
- Pseudo-nitzschia turgidula
References
- ↑ Photo taken by Dr. Rozalind Jester
- 1 2 Maldonado, Maria T. (2002). "The effect of Fe and Cu on growth and domoic acid production by Pseudo-nitzschia multiseries and Pseudo-nitzschia australis". Limnology and Oceanography. 47: 515–526. doi:10.4319/lo.2002.47.2.0515.
- ↑ Lim, Hong-Chang; Leaw, Chui-Pin; Su, Suriyanti Nyun-Pau; Teng, Sing-Tung; Usup, Gires; Mohammad-Noor, Normawaty; Lundholm, Nina; Kotaki, Yuichi; Lim, Po-Teen (2012-10-01). "Morphology and Molecular Characterization of Pseudo-Nitzschia (bacillariophyceae) from Malaysian Borneo, Including the New Species Pseudo-Nitzschia Circumpora Sp. Nov". Journal of Phycology. 48 (5): 1232–1247. doi:10.1111/j.1529-8817.2012.01213.x. ISSN 1529-8817.
- 1 2 Parsons, Michael L.; Dortch, Quay (2002-03-01). "Sedimentological evidence of an increase in Pseudo-nitzschia (Bacillariophyceae)abundance in response to coastal eutrophication". Limnology and Oceanography. 47 (2): 551–558. doi:10.4319/lo.2002.47.2.0551. ISSN 1939-5590.
- 1 2 3 4 5 6 7 Kuwata, Akira; Jewson, David H. (2015-01-01). Ohtsuka, Susumu; Suzaki, Toshinobu; Horiguchi, Takeo; Suzuki, Noritoshi; Not, Fabrice, eds. Ecology and Evolution of Marine Diatoms and Parmales. Springer Japan. pp. 251–275. doi:10.1007/978-4-431-55130-0_10#page-1. ISBN 9784431551294.
- 1 2 Lundholm, Nina; Moestrup, Øjvind. "The marine diatom Pseudo-nitzschia galaxiae sp. nov. (Bacillariophyceae): morphology and phylogenetic relationships". Phycologia. 41 (6): 594–605. doi:10.2216/i0031-8884-41-6-594.1.
- ↑ Hasle, G.R. 1994. Pseudo-nitzschia as a genus distinct from Nitzschia (Bacillariophyceae). J. Phycol. 30: 1036-1039.
- ↑ Lim, H.-C., Leaw, C.-P., Su, S. N.-P., Teng, S.-T., Usup, G., Mohammad-Noor, N., Lundholm, N., Kotaki, Y. and Lim, P.-T. (2012), MORPHOLOGY AND MOLECULAR CHARACTERIZATION OFPSEUDO-NITZSCHIA (BACILLARIOPHYCEAE) FROM MALAYSIAN BORNEO, INCLUDING THE NEW SPECIES PSEUDO-NITZSCHIA CIRCUMPORA SP. NOV. Journal of Phycology, 48: 1232–1247. doi: 10.1111/j.1529-8817.2012.01213.x
- ↑ Noyer, Charlotte; Abot, Anne; Trouilh, Lidwine; Leberre, Véronique Anton; Dreanno, Catherine (2015-05-01). "Phytochip: Development of a DNA-microarray for rapid and accurate identification of Pseudo-nitzschia spp and other harmful algal species". Journal of Microbiological Methods. 112: 55–66. doi:10.1016/j.mimet.2015.03.002.
- ↑ Hasle, G.R. 1995. Pseudo-nitzschia pungens and P. multiseries (Bacillariophyceae): nomenclatural history, morphology, and distribution. J. Phycol. 31: 428-435.
- ↑ Bates, S.S. 2000. Domoic-acid-producing diatoms: another genus added! J. Phycol. 36: 978-983.
- 1 2 Scalco, Eleonora; Amato, Alberto; Ferrante, Maria Immacolata; Montresor, Marina (2015-10-22). "The sexual phase of the diatom Pseudo-nitzschia multistriata: cytological and time-lapse cinematography characterization". Protoplasma: 1–11. doi:10.1007/s00709-015-0891-5. ISSN 0033-183X.
- 1 2 Scalco, Eleonora; Stec, Krzysztof; Iudicone, Daniele; Ferrante, Maria Immacolata; Montresor, Marina (2014-10-01). "The dynamics of sexual phase in the marine diatom Pseudo-nitzschia multistriata (Bacillariophyceae)". Journal of Phycology. 50 (5): 817–828. doi:10.1111/jpy.12225. ISSN 1529-8817.
- ↑ "Info - Pseudo-nitzschia multiseries CLN-47". genome.jgi.doe.gov. Retrieved 2016-05-06.
- ↑ Dato, Valeria Di, Francesco Musacchia, Giuseppe Petrosino, Shrikant Patil, Marina Montresor, Remo Sanges, and Maria Immacolata Ferrante. "Transcriptome Sequencing of Three Pseudo-nitzschia Species Reveals Comparable Gene Sets and the Presence of Nitric Oxide Synthase Genes in Diatoms." Sci. Rep. Scientific Reports 5 (2015): 12329. Web.
- ↑ "NOAA Coastal Science" (PDF).
- 1 2 Dhar, Bidhan Chandra; Cimarelli, Lucia; Singh, Kumar Saurabh; Brandi, Letizia; Brandi, Anna; Puccinelli, Camilla; Marcheggiani, Stefania; Spurio, Roberto (2015-05-06). "Molecular Detection of a Potentially Toxic Diatom Species". International Journal of Environmental Research and Public Health. 12 (5): 4921–4941. doi:10.3390/ijerph120504921. PMC 4454946
 . PMID 25955528.
. PMID 25955528. - ↑ Delegrange, A.; Vincent, D.; Courcot, L.; Amara, R. (2015-02-01). "Testing the vulnerability of juvenile sea bass (Dicentrarchus labrax) exposed to the harmful algal bloom (HAB) species Pseudo-nitzschia delicatissima". Aquaculture. 437: 167–174. doi:10.1016/j.aquaculture.2014.11.023.
- ↑ Bates, Stephen (1998). "Bloom dynamics and physiology of domoic-acid-producing Pseudo-nitzschia species". Physiological Ecology of Harmfull Algal Blooms.
- 1 2 3 Auro, Maureen E.; Cochlan, William P. (2013-02-01). "Nitrogen Utilization and Toxin Production by Two Diatoms of the Pseudo-nitzschia pseudodelicatissima Complex: P. cuspidata and P. fryxelliana". Journal of Phycology. 49 (1): 156–169. doi:10.1111/jpy.12033. ISSN 1529-8817.
- ↑ Martin-Jézéquel, Véronique; Calu, Guillaume; Candela, Leo; Amzil, Zouher; Jauffrais, Thierry; Séchet, Véronique; Weigel, Pierre (2015-11-26). "Effects of Organic and Inorganic Nitrogen on the Growth and Production of Domoic Acid by Pseudo-nitzschia multiseries and P. australis (Bacillariophyceae) in Culture". Marine Drugs. 13 (12): 7067–7086. doi:10.3390/md13127055. PMC 4699229. PMID 26703627.
- ↑ Tammilehto, Anna; Nielsen, Torkel Gissel; Krock, Bernd; Møller, Eva Friis; Lundholm, Nina (2015-02-01). "Induction of domoic acid production in the toxic diatom Pseudo-nitzschia seriata by calanoid copepods". Aquatic Toxicology. 159: 52–61. doi:10.1016/j.aquatox.2014.11.026.
External links
- Domoic Acid and Pseudo-nitzschia References
- IOC-UNESCO HAB Taxon list
- NOAA Phytoplankton Monitoring Network
- Northwest Fisheries Science Center, NOAA
- Toxic Blooms: Understanding Red Tides, a seminar by the Woods Hole Oceanographic Institution
- NOAA Marine Biotoxins Program, National Oceanic and Atmospheric Administration
- California Program for Regional Enhanced Monitoring for PhycoToxins, California Department of Health Services and the University of California, Santa Cruz
- Washington State Shellfish Biotoxin Program
- Series in Marine Phytoplankton: The genus Pseudo-nitzschia