Molybdopterin
 | |
| Identifiers | |
|---|---|
| 73508-07-3 | |
| MeSH | molybdopterin |
| PubChem | 459 |
| Properties | |
| C 10H 10N 5O 6PS 2 + R groups | |
| Molar mass | 394.33 g/mol (R=H) |
| Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
| | |
| Infobox references | |
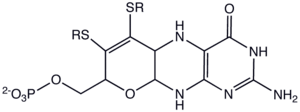
Molybdopterins are a class of cofactors found in most molybdenum (Mo) and all tungsten (W) enzymes. Synonyms for molydopterin are: MPT and pyranopterin-dithiolate. The nomenclature for this biomolecule can be confusing: Molybdopterin per se contains no molybdenum; rather, this is the name of the ligand (a pterin) that will bind the active metal. After molydopterin is eventually complexed with molybdenum, the complete ligand is usually called molybdenum cofactor.
Molydopterin consists of a pyranopterin, a complex heterocycle featuring a pyran fused to a pterin ring. In addition, the pyran ring features two thiolates, which serve as ligands in molybdo- and tungstoenzymes. In some cases, the alkyl phosphate group is replaced by an alkyl diphosphate nucleotide. Enzymes that contain the molybdopterin cofactor include xanthine oxidase, DMSO reductase, sulfite oxidase, and nitrate reductase.
The only molybdenum-containing enzymes that do not feature molydopterins are the nitrogenases (enzymes that fix nitrogen). These contain an iron-sulfur center of a very different type, which also usually contains molybdenum. However, if molybdenum is present, it is directly bonded to other metal atoms.[1]
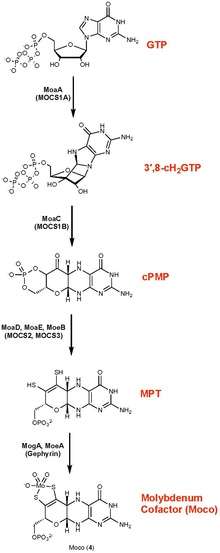
Biosynthesis
The biosynthesis of molybdopterin begins with guanosine triphosphate. Two enzymatic reactions convert this triphosphate to the cyclic phosphate of pyranopterin. One of these enzymes uses the radical SAM mechanism, often associated with C—X bond-forming reactions.[2][3] This intermediate pyranopterin is then converted to the molybdopterin via the action of three further enzymes. In this conversion, the enedithiolate is formed, although the substituents on sulfur remain unknown. Sulfur is conveyed from cysteinyl persulfide in a manner reminiscent of the biosynthesis of iron-sulfur proteins. The monophosphate is adenylated (coupled to ADP) in a step that activates the cofactor toward binding Mo or W. These metals are imported as their oxyanions, molybdate, and tungstate. Finally, Mo or W is inserted to give the molybdopterin cofactor. In some enzymes, such as xanthine oxidase, the metal is bound to one molybdopterin, whereas, in other enzymes, e.g., DMSO reductase, the metal is bound to two molybdopterin cofactors.[4]
Models for the active sites of enzymes molybdopterin-containing enzymes are based on a class of ligands known as dithiolenes.[5]
Tungsten derivatives
Some oxidoreductases use tungsten in a similar manner as molybdenum by using it in a tungsten-pterin complex, with molybdopterin. Thus, molybdopterin may complex with either molybdenum or tungsten, for use by bacteria. Tungsten-using enzymes typically reduce free carboxylic acids to aldehydes.[6]
The first tungsten-requiring enzyme to be discovered also requires selenium (though the precise form is unknown). In this case, the tungsten-selenium pair has been speculated to function analogously to the molybdenum-sulfur pairing of some molybdenum cofactor-requiring enzymes.[7] Although a tungsten-containing xanthine dehydrogenase from bacteria has been found to contain tungsten-molydopterin and also non-protein-bound selenium (thus removing the possibility of selenium in selenocysteine or selenomethionine form), a tungsten-selenium molybdopterin complex has not been definitively described.[8]
Enzymes that use molybdopterin
Enzymes that use molydopterin as cofactor or prosthetic group are given below.[1] Molydopterin is a:
- Cofactor of: xanthine oxidase, DMSO reductase, sulfite oxidase, nitrate reductase, ethylbenzene dehydrogenase, glyceraldehyde-3-phosphate ferredoxin oxidoreductase, respiratory arsenate reductase, carbon monoxide dehydrogenase.
- Prosthetic group of: formate dehydrogenase, purine hydroxylase, thiosulfate reductase.
References
- 1 2 Structure, synthesis, empirical formula for the di-sulfhydryl. Accessed Nov. 16, 2009.
- ↑ Hover BM, Loksztejn A, Ribeiro AA, Yokoyama K (April 2013). "Identification of a cyclic nucleotide as a cryptic intermediate in molybdenum cofactor biosynthesis". J Am Chem Soc. 135 (18): 7019–32. doi:10.1021/ja401781t. PMID 23627491.
- ↑ Hover BM, Tonthat NK, Schumacher MA, Yokoyama K (May 2015). "Mechanism of pyranopterin ring formation in molybdenum cofactor biosynthesis". Proc Natl Acad Sci USA. 112: 6347–52. doi:10.1073/pnas.1500697112. PMID 25941396.
- ↑ Schwarz, G. & Mendel, R. R. (2006). "Molybdenum cofactor biosynthesis and molybdenum enzymes". Annual Review of Plant Biology. 57: 623–647. doi:10.1146/annurev.arplant.57.032905.105437. PMID 16669776.
- ↑ Kisker, C.; Schindelin, H.; Baas, D.; Rétey, J.; Meckenstock, R.U.; Kroneck, P.M.H. (1999). "A structural comparison of molybdenum cofactor-containing enzymes". FEMS Microbiol. Rev. 22 (5): 503–521. doi:10.1111/j.1574-6976.1998.tb00384.x. PMID 9990727.
- ↑ Lassner, Erik (1999). Tungsten: Properties, Chemistry, Technology of the Element, Alloys and Chemical Compounds. Springer. pp. 409–411. ISBN 0-306-45053-4.
- ↑ Stiefel, E. I. (1998). "Transition metal sulfur chemistry and its relevance to molybdenum and tungsten enzymes" (PDF). Pure & Appl. Chem. 70 (4): 889–896. doi:10.1351/pac199870040889.
- ↑ Schräder T, Rienhöfer A, Andreesen JR (September 1999). "Selenium-containing xanthine dehydrogenase from Eubacterium barkeri". Eur. J. Biochem. 264 (3): 862–71. doi:10.1046/j.1432-1327.1999.00678.x. PMID 10491134.