Major facilitator superfamily
| Major Facilitator Superfamily | |
|---|---|
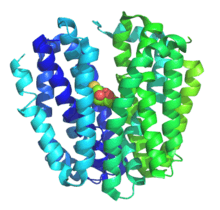 Crystal Structure of Lactose Permease LacY. | |
| Identifiers | |
| Symbol | MFS |
| Pfam clan | CL0015 |
| TCDB | 2.A.1 |
| OPM superfamily | 15 |
The major facilitator superfamily (MFS) is a class of membrane transport proteins that facilitate movement of small solutes across cell membranes in response to chemiosmotic gradients.[1][2]
Function
The major facilitator superfamily (MFS) are membrane proteins which are expressed ubiquitously in all kingdoms of life for the import or export of target substrates. The MFS family was originally believed to function primarily in the uptake of sugars but subsequent studies revealed that drugs, metabolites, oligosaccharides, amino acids and oxyanions were all transported by MFS family members.[3] These protein energetically drive transport utilizing the electrochemical gradient of the target substrate (uniporter), or act as a cotransporter where transport is coupled to the movement of a second substrate.
Fold
The basic fold of the MFS transporter is built around 12 transmembrane helices (TMH),[4] with two 6-helix bundles formed by the N and C terminal homologus domains[5] of the transporter which are connected by an extended cytoplasmic loop. The two halves of the protein pack against each other in a clam-shell fashion, sealing via interactions at the ends of the transmembrane helices and extracellular loops.[6][7] This forms a large aqueous cavity at the center of the membrane, which is alternatively open to the cytoplasm or periplasm/extracellular space. Lining this aqueous cavity are the amino-acids which bind the substrate(s) and define transporter specificity.[8][9] Many MFS transporters are thought to be dimers through in vitro and in vivo methods, with some evidence to suggest a functional role for this oligomerization.[10]
Mechanism
The alternating-access mechanism thought to underlie the transport of most MFS transport is classically described as the "rocker-switch" mechanism.[6][7] In this model, the transporter opens to either the extracellular space or cytoplasm and simultaneously seals the opposing face of the transporter, preventing a continuous pathway across the membrane. For example, in the best studied MFS transporter, LacY, lactose and protons typically bind from the periplasm to specific sites within the aqueous cleft. This drives closure of the extracellular face, and opening of the cytoplasmic side, allowing substrate into the cell. Upon substrate release, the transporter recycles to the periplasmic facing orientation.
Exporters and antiporters of the MFS family follow a similar reaction cycle, though exporters bind substrate in the cytoplasm and extrude it to the extracellular or periplasmic space, while antiporters bind substrate in both states to drive each conformational change. While most MFS structures suggest large, rigid body structural changes with substrate binding, the movements may be small in the cases of small substrates, such as the nitrate transporter NarK.[11]
Transport Reaction
The generalized transport reactions catalyzed by MFS porters are:
(1) Uniport: S (out) ⇌ S (in)
(2) Symport: S (out) + [H+ or Na+] (out) ⇌ S (in) + [H+ or Na+] (in)
(3) Antiport: S1 (out) + S2 (in) ⇌ S1 (in) + S2 (out) (S1 may be H+ or a solute)
Substrate specificity
Though initially identified as sugar transporters, a function conserved from prokaryotes[9] to mammals,[12] the MFS family is notable for the great diversity of substrates transported by the superfamily. These range from small oxyanions[13][14][15] to large peptide fragments.[16] Other MFS transporters are notable for a lack of selectivity, extruding broad classes of drugs and xenobiotics.[17][18][19] This substrate specificity is largely determined by specific side chains which line the aqueous pocket at the center of the membrane.[8][9] While one substrate of particular biological importance is often used to name the transporter or family, there may also be co-transported or leaked ions or molecules. These include water molecules[20][21] or the coupling ion(s) which energetically drive transport.
Structures
The crystal structures of a number of MFS transporters have been characterized. The first structures were of the glycerol 3-phosphate/phosphate exchanger GlpT[7] and the lactose-proton symporter LacY,[6] which served to elucidate the overall structure of the protein family and provided initial models for understanding the MFS transport mechanism. Since these initial structures other MFS structures have been solved which illustrate substrate specificity or states within the reaction cycle.[22][23] While the initial MFS structures solved were of bacterial transporters, recently structures of the first eukaryotic structures have been published. These include a fungal phosphate transporter PiPT,[15] plant nitrate transporter NRT1.1,[10][24] and the human glucose transporter GLUT1.[25]
Evolution
The origin of the basic MFS transporter fold is currently under heavy debate. All currently recognized MFS permeases have the two six-TMH domains within a single polypeptide chain, although in some MFS families an additional two TMHs are present. Evidence suggests that the MFS permeases arose by a tandem intragenic duplication event in the early prokaryotes. This event generated the 12 transmembrane helix topology from a (presumed) primordial 6-helix dimer. Moreover, the well-conserved MFS specific motif between TMS2 and TMS3 and the related but less well conserved motif between TMS8 and TMS9 prove to be a characteristic of virtually all of the more than 300 MFS proteins identified.[26] However, the origin of the primordial 6-helix domain is under heavy debate. While some functional and structural evidence suggests that this domain arose out of a simpler 3-helix domain,[27][28] bioinformatic or phylogenetic evidence supporting this hypothesis is lacking.[29][30]
Medical significance
MFS family members are central to human physiology and play an important role in a number of diseases, through aberrant action, drug transport, or drug resistance. The OAT1 transporter transports a number of nucleoside analogs central to antiviral therapy.[31] Resistance to antibiotics is frequently the result of action of MFS resistance genes.[32] Mutations in MFS transporters have also been found to be cause neurodegerative disease,[33] vascular disorders of the brain,[34] and glucose storage diseases.[35]
Disease mutations
Disease associated mutations have been found in a number of human MFS transporters; those annotated in Uniprot are listed below.
| Name | Uniprot ID | Function | Disease |
|---|---|---|---|
| SLC37A4 | O43826 | Transports glucose-6-phosphate from the cytoplasm to the lumen of the endoplasmic reticulum. Forms with glucose-6-phosphatase the complex responsible for glucose production through glycogenolysis and gluconeogenesis. Hence, it plays a central role in homeostatic regulation of blood glucose levels. | Glycogen storage disease type I[36] |
| FLVCR1 | Q9Y5Y0 | Heme transporter that exports cytoplasmic heme. It can also export coproporphyrin and protoporphyrin IX, which are both intermediate products in the heme biosynthetic pathway. Does not export bilirubin. Heme export depends on the presence of HPX and may be required to protect developing erythroid cells from heme toxicity. Heme export also provides protection from heme or ferrous iron toxicities in liver and brain. Causes susceptibility to FeLV-C in vitro. Required during erythtopoiesis to maintain intracellular free heme balance since in proerythroblasts, heme synthesis intensifies and it's accumulation is toxic for cells. | Retinitis pigmentosa[37] |
| SLC33A1 | O00400 | Probable acetyl-CoA transporter necessary for O-acetylation of gangliosides. | Spastic paraplegia[38] |
| SLC17A5 | Q9NRA2 | Transports glucuronic acid and free sialic acid out of the lysosome after it is cleaved from sialoglycoconjugates undergoing degradation, this is required for normal CNS myelination. Mediates aspartate and glutamate membrane potential-dependent uptake into synaptic vesicles and synaptic-like microvesicles. Also functions as an electrogenic 2NO(3)(-)/H(+) cotransporter in the plasma membrane of salivary gland acinar cells, mediating the physiological nitrate efflux, 25% of the circulating nitrate ions is typically removed and secreted in saliva. | Salla disease[39] |
| SLC2A10 | O95528 | Facilitative glucose transporter. | Arterial tortuosity syndrome[40] |
| SLC22A12 | Q96S37 | Required for efficient urate re-absorption in the kidney. Regulates blood urate levels. Mediates saturable urate uptake by facilitating the exchange of urate against organic anions. | Hypouricemia[41] |
| SLC16A1 | P53985 | Proton-coupled monocarboxylate transporter. Catalyzes the rapid transport across the plasma membrane of many monocarboxylates such as lactate, pyruvate, branched-chain oxo acids derived from leucine, valine and isoleucine, and the ketone bodies acetoacetate, beta-hydroxybutyrate and acetate. Depending on the tissue and on circumstances, mediates the import or export of lactic acid and ketone bodies. Required for normal nutrient assimilation, increase of white adipose tissue and body weight gain when on a high-fat diet. Plays a role in cellular responses to a high-fat diet by modulating the cellular levels of lactate and pyruvate, small molecules that contribute to the regulation of central metabolic pathways and insulin secretion, with concomitant effects on plasma insulin levels and blood glucose homeostasis. | Hypoglycemia[42] |
| SLC22A5 | O76082 | Sodium-ion dependent, high affinity carnitine transporter. Involved in the active cellular uptake of carnitine. Transports one sodium ion with one molecule of carnitine. Also transports organic cations such as tetraethylammonium (TEA) without the involvement of sodium. | Systemic primary carnitine deficiency[43] |
| CLN3 | Q13286 | Involved in microtubule-dependent, anterograde transport of late endosomes and lysosomes. | Ceroid lipofuscinosis[44] |
| SLC16A13 | Q7RTY0 | Proton-linked monocarboxylate transporter. Catalyzes the rapid transport across the plasma membrane of many monocarboxylates (By similarity). | Diabetes mellitus[45] |
| SLC2A9 | Q9NRM0 | Transport urate and fructose. May have a role in the urate reabsorption by proximal tubules. Also transports glucose at low rate. | Hypouricemia[46] |
| SLC19A3 | Q9BZV2 | Mediates high affinity thiamine uptake, probably via a proton anti-port mechanism. | Thiamine metabolism dysfunction syndrome[47] |
| FLVCR2 | Q9UPI3 | Acts as an importer of heme. Also acts as a transporter for a calcium-chelator complex, important for growth and calcium metabolism. | Fowler syndrome[34] |
| SLC16A12 | Q6ZSM3 | Proton-linked monocarboxylate transporter. Catalyzes the rapid transport across the plasma membrane of many monocarboxylates (By similarity). | Cataract[48] |
| SLC19A2 | O60779 | High-affinity transporter for the intake of thiamine. | Megaloblastic Anemia[49] |
| MFSD8 | Q8NHS3 | May be a carrier that transport small solutes by using chemiosmotic ion gradients (Potential). | Ceroid lipofuscinosis[50] |
| SLC40A1 | Q9NP59 | May be involved in iron export from duodenal epithelial cell and also in transfer of iron between maternal and fetal circulation. Mediates iron efflux in the presence of a ferroxidase (hephaestin and/or ceruloplasmin). | Hemochromatosis[51] |
| SLC2A4 | P14672 | Insulin-regulated facilitative glucose transporter. | Diabetes mellitus[52] |
| SLC45A2 | Q9UMX9 | Melanocyte differentiation antigen. May transport substances required for melanin biosynthesis (By similarity). | Albinism[53] |
| SLCO2A1 | Q92959 | May mediate the release of newly synthesized prostaglandins from cells, the transepithelial transport of prostaglandins, and the clearance of prostaglandins from the circulation. Transports PGD2, as well as PGE1, PGE2 and PGF2A. | Hypertrophic osteoarthropathy[54] |
| SLC22A4 | Q9H015 | Sodium-ion dependent, low affinity carnitine transporter. Probably transports one sodium ion with one molecule of carnitine. Also transports organic cations such as tetraethylammonium (TEA) without the involvement of sodium. Relative uptake activity ratio of carnitine to TEA is 1.78. A key substrate of this transporter seems to be ergothioneine (ET). | Rheumatoid arthritis[55] |
| SLC16A11 | Q8NCK7 | Proton-linked monocarboxylate transporter. Catalyzes the rapid transport across the plasma membrane of many monocarboxylates (By similarity). Probably involved in hepatic lipid metabolism: overexpression results in an increase of triacylglycerol(TAG) levels, small increases in intracellular diacylglycerols and decreases in lysophosphatidylcholine, cholesterol ester and sphingomyelin lipids. | Diabetes mellitus[45] |
| SLCO1B3 | Q9NPD5 | Mediates the Na(+)-independent uptake of organic anions such as 17-beta-glucuronosyl estradiol, taurocholate, triiodothyronine (T3), leukotriene C4, dehydroepiandrosterone sulfate (DHEAS), methotrexate and sulfobromophthalein (BSP). Involved in the clearance of bile acids and organic anions from the liver. | Hyperbilirubinemia[56] |
| SLCO1B1 | Q9Y6L6 | Mediates the Na(+)-independent uptake of organic anions such as pravastatin, taurocholate, methotrexate, dehydroepiandrosterone sulfate, 17-beta-glucuronosyl estradiol, estrone sulfate, prostaglandin E2, thromboxane B2, leukotriene C3, leukotriene E4, thyroxine and triiodothyronine. Involved in the clearance of bile acids and organic anions from the liver. | Hyperbilirubinemia[56] |
| SLC2A2 | P11168 | Facilitative glucose transporter. This isoform likely mediates the bidirectional transfer of glucose across the plasma membrane of hepatocytes and is responsible for uptake of glucose by the beta cells; may comprise part of the glucose-sensing mechanism of the beta cell. May also participate with the Na(+)/glucose cotransporter in the transcellular transport of glucose in the small intestine and kidney. | Fanconi-Bickel syndrome[57] |
| SLC2A1 | P11166 | Facilitative glucose transporter. This isoform may be responsible for constitutive or basal glucose uptake. Has a very broad substrate specificity; can transport a wide range of aldoses including both pentoses and hexoses. | GLUT1 deficiency syndrome 1[58] |
| SLC46A1 | Q96NT5 | Has been shown to act both as an intestinal proton-coupled high-affinity folate transporter and as an intestinal heme transporter which mediates heme uptake from the gut lumen into duodenal epithelial cells. The iron is then released from heme and may be transported into the bloodstream. Dietary heme iron is an important nutritional source of iron. Shows a higher affinity for folate than heme. | Hereditary folate malabsorption[59] |
| SLC17A8 | Q8NDX2 | Mediates the uptake of glutamate into synaptic vesicles at presynaptic nerve terminals of excitatory neural cells. May also mediate the transport of inorganic phosphate. | Deafness[60] |
References
- ↑ Pao SS, Paulsen IT, Saier MH (1998). "Major Facilitator Superfamily". Microbiol. Mol. Biol. Rev. 62 (1): 1–34. PMC 98904
 . PMID 9529885.
. PMID 9529885. - ↑ Walmsley AR, Barrett MP, Bringaud F, Gould GW (December 1998). "Sugar transporters from bacteria, parasites and mammals: structure-activity relationships". Trends Biochem. Sci. 23 (12): 476–81. doi:10.1016/S0968-0004(98)01326-7. PMID 9868370.
- ↑ Marger MD, Saier MH (Jan 1993). "A major superfamily of transmembrane facilitators that catalyze uniport, symport and antiport.". Trends in Biochemical Sciences. 18 (1): 13–20. doi:10.1016/0968-0004(93)90081-w. PMID 8438231.
- ↑ Foster DL, Boublik M, Kaback HR (Jan 10, 1983). "Structure of the lac carrier protein of Escherichia coli.". The Journal of Biological Chemistry. 258 (1): 31–4. PMID 6336750.
- ↑ Maiden MC, Davis EO, Baldwin SA, Moore DC, Henderson PJ (Feb 12–18, 1987). "Mammalian and bacterial sugar transport proteins are homologous.". Nature. 325 (6105): 641–3. doi:10.1038/325641a0. PMID 3543693.
- 1 2 3 Abramson J, Smirnova I, Kasho V, Verner G, Kaback HR, Iwata S (Aug 1, 2003). "Structure and mechanism of the lactose permease of Escherichia coli.". Science. 301 (5633): 610–5. doi:10.1126/science.1088196. PMID 12893935.
- 1 2 3 Huang Y, Lemieux MJ, Song J, Auer M, Wang DN (Aug 1, 2003). "Structure and mechanism of the glycerol-3-phosphate transporter from Escherichia coli.". Science. 301 (5633): 616–20. doi:10.1126/science.1087619. PMID 12893936.
- 1 2 Yan N (Mar 2013). "Structural advances for the major facilitator superfamily (MFS) transporters.". Trends in Biochemical Sciences. 38 (3): 151–9. doi:10.1016/j.tibs.2013.01.003. PMID 23403214.
- 1 2 3 Kaback HR, Sahin-Tóth M, Weinglass AB (Aug 2001). "The kamikaze approach to membrane transport.". Nature reviews. Molecular cell biology. 2 (8): 610–20. doi:10.1038/35085077. PMID 11483994.
- 1 2 Sun J, Bankston JR, Payandeh J, Hinds TR, Zagotta WN, Zheng N (Mar 6, 2014). "Crystal structure of the plant dual-affinity nitrate transporter NRT1.1.". Nature. 507 (7490): 73–7. doi:10.1038/nature13074. PMID 24572362.
- ↑ Zheng H, Wisedchaisri G, Gonen T (May 30, 2013). "Crystal structure of a nitrate/nitrite exchanger.". Nature. 497 (7451): 647–51. doi:10.1038/nature12139. PMID 23665960.
- ↑ Mueckler M, Caruso C, Baldwin SA, Panico M, Blench I, Morris HR, Allard WJ, Lienhard GE, Lodish HF (Sep 6, 1985). "Sequence and structure of a human glucose transporter.". Science. 229 (4717): 941–5. doi:10.1126/science.3839598. PMID 3839598.
- ↑ Yan H, Huang W, Yan C, Gong X, Jiang S, Zhao Y, Wang J, Shi Y (Mar 28, 2013). "Structure and mechanism of a nitrate transporter.". Cell reports. 3 (3): 716–23. doi:10.1016/j.celrep.2013.03.007. PMID 23523348.
- ↑ Tsay YF, Schroeder JI, Feldmann KA, Crawford NM (Mar 12, 1993). "The herbicide sensitivity gene CHL1 of Arabidopsis encodes a nitrate-inducible nitrate transporter.". Cell. 72 (5): 705–13. doi:10.1016/0092-8674(93)90399-b. PMID 8453665.
- 1 2 Pedersen BP, Kumar H, Waight AB, Risenmay AJ, Roe-Zurz Z, Chau BH, Schlessinger A, Bonomi M, Harries W, Sali A, Johri AK, Stroud RM (Apr 25, 2013). "Crystal structure of a eukaryotic phosphate transporter.". Nature. 496 (7446): 533–6. doi:10.1038/nature12042. PMID 23542591.
- ↑ Doki S, Kato HE, Solcan N, Iwaki M, Koyama M, Hattori M, Iwase N, Tsukazaki T, Sugita Y, Kandori H, Newstead S, Ishitani R, Nureki O (Jul 9, 2013). "Structural basis for dynamic mechanism of proton-coupled symport by the peptide transporter POT.". Proceedings of the National Academy of Sciences of the United States of America. 110 (28): 11343–8. doi:10.1073/pnas.1301079110. PMID 23798427.
- ↑ Jiang D, Zhao Y, Wang X, Fan J, Heng J, Liu X, Feng W, Kang X, Huang B, Liu J, Zhang XC (Sep 3, 2013). "Structure of the YajR transporter suggests a transport mechanism based on the conserved motif A.". Proceedings of the National Academy of Sciences of the United States of America. 110 (36): 14664–9. doi:10.1073/pnas.1308127110. PMID 23950222.
- ↑ Putman M, van Veen HW, Konings WN (Dec 2000). "Molecular properties of bacterial multidrug transporters.". Microbiology and molecular biology reviews : MMBR. 64 (4): 672–93. doi:10.1128/mmbr.64.4.672-693.2000. PMID 11104814.
- ↑ Yin Y, He X, Szewczyk P, Nguyen T, Chang G (May 5, 2006). "Structure of the multidrug transporter EmrD from Escherichia coli.". Science. 312 (5774): 741–4. doi:10.1126/science.1125629. PMID 16675700.
- ↑ Li J, Shaikh SA, Enkavi G, Wen PC, Huang Z, Tajkhorshid E (May 7, 2013). "Transient formation of water-conducting states in membrane transporters.". Proceedings of the National Academy of Sciences of the United States of America. 110 (19): 7696–701. doi:10.1073/pnas.1218986110. PMID 23610412.
- ↑ Fischbarg J, Kuang KY, Vera JC, Arant S, Silverstein SC, Loike J, Rosen OM (Apr 1990). "Glucose transporters serve as water channels.". Proceedings of the National Academy of Sciences of the United States of America. 87 (8): 3244–7. doi:10.1073/pnas.87.8.3244. PMID 2326282.
- ↑ Dang S, Sun L, Huang Y, Lu F, Liu Y, Gong H, Wang J, Yan N (Oct 7, 2010). "Structure of a fucose transporter in an outward-open conformation.". Nature. 467 (7316): 734–8. doi:10.1038/nature09406. PMID 20877283.
- ↑ Kumar H, Kasho V, Smirnova I, Finer-Moore JS, Kaback HR, Stroud RM (Feb 4, 2014). "Structure of sugar-bound LacY.". Proceedings of the National Academy of Sciences of the United States of America. 111 (5): 1784–8. doi:10.1073/pnas.1324141111. PMID 24453216.
- ↑ Parker JL, Newstead S (Mar 6, 2014). "Molecular basis of nitrate uptake by the plant nitrate transporter NRT1.1.". Nature. 507 (7490): 68–72. doi:10.1038/nature13116. PMID 24572366.
- ↑ Deng D, Xu C, Sun P, Wu J, Yan C, Hu M, Yan N (Jun 5, 2014). "Crystal structure of the human glucose transporter GLUT1.". Nature. 510 (7503): 121–5. doi:10.1038/nature13306. PMID 24847886.
- ↑ Henderson PJ (Mar–Apr 1990). "The homologous glucose transport proteins of prokaryotes and eukaryotes.". Research in microbiology. 141 (3): 316–28. doi:10.1016/0923-2508(90)90005-b. PMID 2177911.
- ↑ Madej MG, Dang S, Yan N, Kaback HR (Apr 9, 2013). "Evolutionary mix-and-match with MFS transporters.". Proceedings of the National Academy of Sciences of the United States of America. 110 (15): 5870–4. doi:10.1073/pnas.1303538110. PMID 23530251.
- ↑ Madej MG, Kaback HR (Dec 10, 2013). "Evolutionary mix-and-match with MFS transporters II.". Proceedings of the National Academy of Sciences of the United States of America. 110 (50): E4831–8. doi:10.1073/pnas.1319754110. PMID 24259711.
- ↑ Västermark A, Lunt B, Saier M (2014). "Major Facilitator Superfamily Porters, LacY, FucP and XylE of Escherichia coli Appear to Have Evolved Positionally Dissimilar Catalytic Residues without Rearrangement of 3-TMS Repeat Units.". Journal of molecular microbiology and biotechnology. 24 (2): 82–90. doi:10.1159/000358429. PMID 24603210.
- ↑ Västermark A, Saier MH (Apr 1, 2014). "Major Facilitator Superfamily (MFS) evolved without 3-transmembrane segment unit rearrangements.". Proceedings of the National Academy of Sciences of the United States of America. 111 (13): E1162–3. doi:10.1073/pnas.1400016111. PMID 24567407.
- ↑ Wada S, Tsuda M, Sekine T, Cha SH, Kimura M, Kanai Y, Endou H (2000). "Rat multispecific organic anion transporter 1 (rOAT1) transports zidovudine, acyclovir, and other antiviral nucleoside analogs". J. Pharmacol. Exp. Ther. 294 (3): 844–9. PMID 10945832.
- ↑ Fluman N, Bibi E (May 2009). "Bacterial multidrug transport through the lens of the major facilitator superfamily.". Biochimica et Biophysica Acta. 1794 (5): 738–47. doi:10.1016/j.bbapap.2008.11.020. PMID 19103310.
- ↑ Aldahmesh MA, Al-Hassnan ZN, Aldosari M, Alkuraya FS (Oct 2009). "Neuronal ceroid lipofuscinosis caused by MFSD8 mutations: a common theme emerging.". Neurogenetics. 10 (4): 307–11. doi:10.1007/s10048-009-0185-1. PMID 19277732.
- 1 2 Meyer E, Ricketts C, Morgan NV, Morris MR, Pasha S, Tee LJ, Rahman F, Bazin A, Bessières B, Déchelotte P, Yacoubi MT, Al-Adnani M, Marton T, Tannahill D, Trembath RC, Fallet-Bianco C, Cox P, Williams D, Maher ER (Mar 12, 2010). "Mutations in FLVCR2 are associated with proliferative vasculopathy and hydranencephaly-hydrocephaly syndrome (Fowler syndrome).". American Journal of Human Genetics. 86 (3): 471–8. doi:10.1016/j.ajhg.2010.02.004. PMID 20206334.
- ↑ Pascual JM, Wang D, Lecumberri B, Yang H, Mao X, Yang R, De Vivo DC (May 2004). "GLUT1 deficiency and other glucose transporter diseases.". European Journal of Endocrinology. 150 (5): 627–33. doi:10.1530/eje.0.1500627. PMID 15132717.
- ↑ Gerin I, Veiga-da-Cunha M, Achouri Y, Collet JF, Van Schaftingen E (Dec 15, 1997). "Sequence of a putative glucose 6-phosphate translocase, mutated in glycogen storage disease type Ib.". FEBS Letters. 419 (2-3): 235–8. doi:10.1016/s0014-5793(97)01463-4. PMID 9428641.
- ↑ Rajadhyaksha AM, Elemento O, Puffenberger EG, Schierberl KC, Xiang JZ, Putorti ML, Berciano J, Poulin C, Brais B, Michaelides M, Weleber RG, Higgins JJ (Nov 12, 2010). "Mutations in FLVCR1 cause posterior column ataxia and retinitis pigmentosa.". American Journal of Human Genetics. 87 (5): 643–54. doi:10.1016/j.ajhg.2010.10.013. PMID 21070897.
- ↑ Lin P, Li J, Liu Q, Mao F, Li J, Qiu R, Hu H, Song Y, Yang Y, Gao G, Yan C, Yang W, Shao C, Gong Y (Dec 2008). "A missense mutation in SLC33A1, which encodes the acetyl-CoA transporter, causes autosomal-dominant spastic paraplegia (SPG42).". American Journal of Human Genetics. 83 (6): 752–9. doi:10.1016/j.ajhg.2008.11.003. PMC 2668077. PMID 19061983.
- ↑ Verheijen FW, Verbeek E, Aula N, Beerens CE, Havelaar AC, Joosse M, Peltonen L, Aula P, Galjaard H, van der Spek PJ, Mancini GM (Dec 1999). "A new gene, encoding an anion transporter, is mutated in sialic acid storage diseases.". Nature Genetics. 23 (4): 462–5. doi:10.1038/70585. PMID 10581036.
- ↑ Coucke PJ, Willaert A, Wessels MW, Callewaert B, Zoppi N, De Backer J, Fox JE, Mancini GM, Kambouris M, Gardella R, Facchetti F, Willems PJ, Forsyth R, Dietz HC, Barlati S, Colombi M, Loeys B, De Paepe A (Apr 2006). "Mutations in the facilitative glucose transporter GLUT10 alter angiogenesis and cause arterial tortuosity syndrome.". Nature Genetics. 38 (4): 452–7. doi:10.1038/ng1764. PMID 16550171.
- ↑ Vázquez-Mellado J, Jiménez-Vaca AL, Cuevas-Covarrubias S, Alvarado-Romano V, Pozo-Molina G, Burgos-Vargas R (Feb 2007). "Molecular analysis of the SLC22A12 (URAT1) gene in patients with primary gout.". Rheumatology (Oxford, England). 46 (2): 215–9. doi:10.1093/rheumatology/kel205. PMID 16837472.
- ↑ Otonkoski T, Jiao H, Kaminen-Ahola N, Tapia-Paez I, Ullah MS, Parton LE, Schuit F, Quintens R, Sipilä I, Mayatepek E, Meissner T, Halestrap AP, Rutter GA, Kere J (Sep 2007). "Physical exercise-induced hypoglycemia caused by failed silencing of monocarboxylate transporter 1 in pancreatic beta cells.". American Journal of Human Genetics. 81 (3): 467–74. doi:10.1086/520960. PMID 17701893.
- ↑ Burwinkel B, Kreuder J, Schweitzer S, Vorgerd M, Gempel K, Gerbitz KD, Kilimann MW (Aug 2, 1999). "Carnitine transporter OCTN2 mutations in systemic primary carnitine deficiency: a novel Arg169Gln mutation and a recurrent Arg282ter mutation associated with an unconventional splicing abnormality.". Biochemical and Biophysical Research Communications. 261 (2): 484–7. doi:10.1006/bbrc.1999.1060. PMID 10425211.
- ↑ Munroe PB, Mitchison HM, O'Rawe AM, Anderson JW, Boustany RM, Lerner TJ, Taschner PE, de Vos N, Breuning MH, Gardiner RM, Mole SE (Aug 1997). "Spectrum of mutations in the Batten disease gene, CLN3.". American Journal of Human Genetics. 61 (2): 310–6. doi:10.1086/514846. PMC 1715900. PMID 9311735.
- 1 2 Williams AL, Jacobs SB, Moreno-Macías H, Huerta-Chagoya A, Churchhouse C, Márquez-Luna C, García-Ortíz H, Gómez-Vázquez MJ, Burtt NP, Aguilar-Salinas CA, González-Villalpando C, Florez JC, Orozco L, Haiman CA, Tusié-Luna T, Altshuler D (Feb 6, 2014). "Sequence variants in SLC16A11 are a common risk factor for type 2 diabetes in Mexico.". Nature. 506 (7486): 97–101. doi:10.1038/nature12828. PMID 24390345.
- ↑ Matsuo H, Chiba T, Nagamori S, Nakayama A, Domoto H, Phetdee K, Wiriyasermkul P, Kikuchi Y, Oda T, Nishiyama J, Nakamura T, Morimoto Y, Kamakura K, Sakurai Y, Nonoyama S, Kanai Y, Shinomiya N (Dec 2008). "Mutations in glucose transporter 9 gene SLC2A9 cause renal hypouricemia.". American Journal of Human Genetics. 83 (6): 744–51. doi:10.1016/j.ajhg.2008.11.001. PMID 19026395.
- ↑ Zeng WQ, Al-Yamani E, Acierno JS, Slaugenhaupt S, Gillis T, MacDonald ME, Ozand PT, Gusella JF (Jul 2005). "Biotin-responsive basal ganglia disease maps to 2q36.3 and is due to mutations in SLC19A3.". American Journal of Human Genetics. 77 (1): 16–26. doi:10.1086/431216. PMC 1226189. PMID 15871139.
- ↑ Kloeckener-Gruissem B, Vandekerckhove K, Nürnberg G, Neidhardt J, Zeitz C, Nürnberg P, Schipper I, Berger W (Mar 2008). "Mutation of solute carrier SLC16A12 associates with a syndrome combining juvenile cataract with microcornea and renal glucosuria.". American Journal of Human Genetics. 82 (3): 772–9. doi:10.1016/j.ajhg.2007.12.013. PMID 18304496.
- ↑ Labay V, Raz T, Baron D, Mandel H, Williams H, Barrett T, Szargel R, McDonald L, Shalata A, Nosaka K, Gregory S, Cohen N (Jul 1999). "Mutations in SLC19A2 cause thiamine-responsive megaloblastic anaemia associated with diabetes mellitus and deafness.". Nature Genetics. 22 (3): 300–4. doi:10.1038/10372. PMID 10391221.
- ↑ Kousi M, Siintola E, Dvorakova L, Vlaskova H, Turnbull J, Topcu M, Yuksel D, Gokben S, Minassian BA, Elleder M, Mole SE, Lehesjoki AE (Mar 2009). "Mutations in CLN7/MFSD8 are a common cause of variant late-infantile neuronal ceroid lipofuscinosis.". Brain : a journal of neurology. 132 (Pt 3): 810–9. doi:10.1093/brain/awn366. PMID 19201763.
- ↑ Zaahl MG, Merryweather-Clarke AT, Kotze MJ, van der Merwe S, Warnich L, Robson KJ (Oct 2004). "Analysis of genes implicated in iron regulation in individuals presenting with primary iron overload.". Human Genetics. 115 (5): 409–17. doi:10.1007/s00439-004-1166-y. PMID 15338274.
- ↑ Kusari J, Verma US, Buse JB, Henry RR, Olefsky JM (Oct 1991). "Analysis of the gene sequences of the insulin receptor and the insulin-sensitive glucose transporter (GLUT-4) in patients with common-type non-insulin-dependent diabetes mellitus.". The Journal of Clinical Investigation. 88 (4): 1323–30. doi:10.1172/JCI115437. PMC 295602. PMID 1918382.
- ↑ Newton JM, Cohen-Barak O, Hagiwara N, Gardner JM, Davisson MT, King RA, Brilliant MH (Nov 2001). "Mutations in the human orthologue of the mouse underwhite gene (uw) underlie a new form of oculocutaneous albinism, OCA4.". American Journal of Human Genetics. 69 (5): 981–8. doi:10.1086/324340. PMC 1274374. PMID 11574907.
- ↑ Seifert W, Kühnisch J, Tüysüz B, Specker C, Brouwers A, Horn D (Apr 2012). "Mutations in the prostaglandin transporter encoding gene SLCO2A1 cause primary hypertrophic osteoarthropathy and isolated digital clubbing.". Human Mutation. 33 (4): 660–4. doi:10.1002/humu.22042. PMID 22331663.
- ↑ Tokuhiro S, Yamada R, Chang X, Suzuki A, Kochi Y, Sawada T, Suzuki M, Nagasaki M, Ohtsuki M, Ono M, Furukawa H, Nagashima M, Yoshino S, Mabuchi A, Sekine A, Saito S, Takahashi A, Tsunoda T, Nakamura Y, Yamamoto K (Dec 2003). "An intronic SNP in a RUNX1 binding site of SLC22A4, encoding an organic cation transporter, is associated with rheumatoid arthritis.". Nature Genetics. 35 (4): 341–8. doi:10.1038/ng1267. PMID 14608356.
- 1 2 van de Steeg E, Stránecký V, Hartmannová H, Nosková L, Hřebíček M, Wagenaar E, van Esch A, de Waart DR, Oude Elferink RP, Kenworthy KE, Sticová E, al-Edreesi M, Knisely AS, Kmoch S, Jirsa M, Schinkel AH (Feb 1, 2012). "Complete OATP1B1 and OATP1B3 deficiency causes human Rotor syndrome by interrupting conjugated bilirubin reuptake into the liver.". The Journal of Clinical Investigation. 122 (2): 519–28. doi:10.1172/JCI59526. PMID 22232210.
- ↑ Sakamoto O, Ogawa E, Ohura T, Igarashi Y, Matsubara Y, Narisawa K, Iinuma K (Nov 2000). "Mutation analysis of the GLUT2 gene in patients with Fanconi-Bickel syndrome.". Pediatric research. 48 (5): 586–9. doi:10.1203/00006450-200011000-00005. PMID 11044475.
- ↑ Wang D, Kranz-Eble P, De Vivo DC (Sep 2000). "Mutational analysis of GLUT1 (SLC2A1) in Glut-1 deficiency syndrome.". Human Mutation. 16 (3): 224–31. doi:10.1002/1098-1004(200009)16:3<224::AID-HUMU5>3.0.CO;2-P. PMID 10980529.
- ↑ Qiu A, Jansen M, Sakaris A, Min SH, Chattopadhyay S, Tsai E, Sandoval C, Zhao R, Akabas MH, Goldman ID (Dec 1, 2006). "Identification of an intestinal folate transporter and the molecular basis for hereditary folate malabsorption.". Cell. 127 (5): 917–28. doi:10.1016/j.cell.2006.09.041. PMID 17129779.
- ↑ Ruel J, Emery S, Nouvian R, Bersot T, Amilhon B, Van Rybroek JM, Rebillard G, Lenoir M, Eybalin M, Delprat B, Sivakumaran TA, Giros B, El Mestikawy S, Moser T, Smith RJ, Lesperance MM, Puel JL (Aug 2008). "Impairment of SLC17A8 encoding vesicular glutamate transporter-3, VGLUT3, underlies nonsyndromic deafness DFNA25 and inner hair cell dysfunction in null mice.". American Journal of Human Genetics. 83 (2): 278–92. doi:10.1016/j.ajhg.2008.07.008. PMC 2495073. PMID 18674745.