LGR5


| View/Edit Human | View/Edit Mouse |
Leucine-rich repeat-containing G-protein coupled receptor 5 (LGR5) also known as G-protein coupled receptor 49 (GPR49) or G-protein coupled receptor 67 (GPR67) is a protein that in humans is encoded by the LGR5 gene.[3][4] It is a member of GPCR class A orphan receptor proteins. LGR5 is expressed across a diverse range of tissue such as in the muscle, placenta, spinal cord and brain and particularly as a biomarker of adult stem cells in certain tissues.[5]
Gene
Prior to its current naming designation, LGR5 was also known as FEX, HG38, GPR49, and GPR67.[6] The Human LGR5 gene is 144,810 bases long and located at chromosome 12 at position 12q22-q23.[6] Both human, rat and mouse homologs contain 907 amino acids and seven transmembrane domains.[7] After translation, the signal peptide (amino acids 1-21) is cleaved off and the mature peptide (amino acids 22-907) inserts its transmembrane domain into the translocon membrane prior to packaging towards the plasma membrane.
Protein Structure
LGR5 is highly conserved within the mammalian clade. Sequence analyses showed that the transmembrane regions and cysteine-flanked junction between TM1 and the extracellular domain were highly conserved in sea anemone (Anthopleura elegantissima), fly (Drosophila melanogaster), worm (Caenorhabditis elegans), snail (Lymnaea stagnalis), rat (Rattus rattus) and human (Homo sapiens).[5] Homology amongst the metazoan suggests that it has been conserved across animals and was hypothesised as a chimeric fusion of an ancestral GPCR and a leucine-rich repeat motif.
Sheau Hsu, Shan Liang and Aaron Hsueh first identified LGR5, together with LGR4, in 1998 at the University Medical School Stanford, California using expression sequence tags based on putative glycoprotein hormone receptors in Drosphila.[5]
Experimental evidence show that the mature receptor protein contains up to 17 leucine-rich repeats, each composed of 24 amino acids spanning the extracellular domain flanked by the cysteine-rich N-terminal and C-terminal regions. In contrast, other glycoprotein hormone receptors such as Luteinizing hormone, Follicle-stimulating hormone and Thyroid-stimulating hormone contain only 9 repeats.[5] Sequence alignment showed that the second N-glycosylation site in LGR5 (Asn 208) aligns with that on the sixth repeat of gonadotropin and TSH receptors. The cysteine residues flanking the ectodomain form stabilising disulfide bonds that support the secondary structure of the leucine-rich repeats.
Function
LGR5 is a member of the Wnt signaling pathway. Although its ligand remains elusive, it has been shown that costimulation with R-spondin 1 and Wnt-3a induce increased internalization of LGR5. LGR5 also cointernalizes with LRP6 and FZD5 via a clathrin-dependent pathway to form a ternary complex upon Wnt ligand binding. Moreover, the rapid cointernalization of LRP6 by LGR5 induces faster rates of degradation for the former. It has been shown that the C-terminal region of LGR5 is crucial for both dynamic internalization and degradation to occur, although C-terminal truncation does not inhibit LRP6 interaction and internalization, but rather, heightens receptor activity. Thus, only the initial interaction with its unknown ligand and other membrane bound receptors is crucial in its role in Wnt signalling and not the internalization itself.[8] LGR5 is crucial during embryogenesis as LGR null studies in mice incurred 100% neonatal mortality accompanied by several craniofacial distortions such as ankyloglossia and gastrointestinal dilation.[9]
Ligand
LGR5 belongs to a class of class A GPCR orphan receptors. Thus its ligands remain elusive. However, it has been shown that Lgr2, the fly orthologue of mammalian LGR5, binds with “high affinity and specificity” with bursicon, an insect heterodimeric, neurohormone that belongs in the same class as FSH, LH and TSH, which in turn are homologous to mammalian bone morphogenetic factors (BMPs) such as gremlin and cerberus. Therefore, LGR5 might be a receptor for a member of the large family of bone morphogenetic protein antagonists.[10] Moreover, R-spondin proteins were shown to interact with the extracellular domain of LRG5 although the molecular mechanism is not yet fully understood.[11]
Clinical Relevance
LGR5 are well-established stem cell markers in certain types of tissue, wholly due to the fact that they are highly enriched in truly, multipotent stem cells compared to their immediate progeny, the transit-amplifying cells.
Intestines
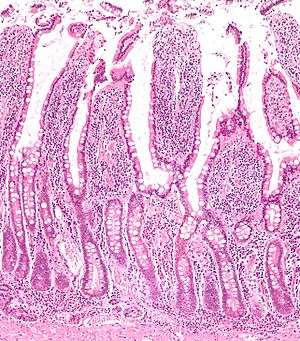
Tracing has revealed that LGR5 is a marker of adult intestinal stem cells. The high turnover rate of the intestinal lining is due to a dedicated population of stem cells found at the base of the intestinal crypt. In the small intestines, these LGR5+ve crypt base columnar cells (CBC cells) have broad basal surfaces and very little cytoplasm and organelles and are located interspersed among the terminally differentiated Paneth cells.[10] These CBC cells generate the plethora of functional cells in the intestinal tissue: Paneth cells, enteroendocrine cells, goblet cells, tuft cells, columnar cells and the M cells over an adult’s entire lifetime. Similarly, LGR5 expression in the colon resembles faithfully that of the small intestine.[10]
Kidney
In vivo lineage tracing showed that LGR5 is expressed in nascent nephron cell cluster within the developing kidney. Specifically, the LGR5+ve stem cells contribute into the formation of the thick ascending limb of Henle’s loop and the distal convoluted tubule. However, expression is eventually truncated after postnatal day 7, a stark contrast to the facultative expression of LGR5 in actively renewing tissues such as in the intestines.[12]
Stomach
The stomach lining also possess populations of LGR5+ve stem cells, although there are two conflicting theories: one is that LGR5+ve stem cells reside in the isthmus, the region between the pit cells and gland cells, where most cellular proliferation takes place. However, lineage tracing had revealed Lgr5+ve stem cells at the bottom of the gland,[13] architecture reminiscent to that of the intestinal arrangement. This suggests that LGR5 stem cells give rise to transit-amplifying cells, which migrate towards the isthmus where they proliferate and maintain the stomach epithelium.[10]
Ear
LGR5+ve stem cells were pinpointed as the precursor for sensory hair cells that line the cochlea (13).
Hair Follicle
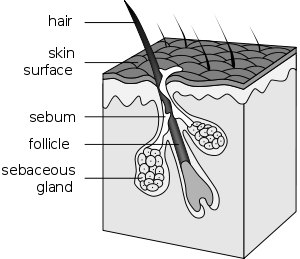
Hair follicle renewal is governed by Wnt signalling that act upon hair follicle stem cells located in the follicle bulge. Although these cells are well characterised by CD34 and cytokeratin markers, there is a growing body of agreement that LGR5 is a putative hair follicle stem cell marker.[14] LGR5 in conjunction with LRG6, is expressed in a remarkable fashion: LRG6+ve stem cells maintain the upper sebaceous gland whilst LRG5+ve stem cells fuel the actual hair follicle shaft upon migration of transit-amplifying cells into the dermal papilla. In between these two distinct populations of stem cells are the multipotent LRG5/6+ve stem cells that ultimately maintain the epidermal hair follicle in adults.[10]
Cancer
The cancer stem cell hypothesis states that a dedicated small population of cancerous stem cells[15] that manages to evade anti-cancer therapy maintains benign and malignant tumours. This explains recurring malignancies even after surgical removal of the tumours.[4] LGR5+ve stem cells were identified to fuel stem cell activity in murine intestinal adenomas via erroneous activation of the pro-cell cycle Wnt signalling pathway as a result of successive mutations, such as formation of adenoma via Adenomatous polyposis coli (APC) mutation.[16] Studies on LGR5 in colorectal cancer revealed a rather perplexing mechanism: loss of LGR5 actually increased tumourigenicity and invasion whereas overexpression results a reduction in tumourigenicity and clonogenicity. This implies that LGR5 is not an oncogene but a tumor suppressor gene, and that its main role is delimiting stem cell expansion in their respective niches.[17] Varying expression profile of LGR5 was also observed in different stages of gastrointestinal cancers, which suggests that the histoanatomical distribution of LGR5+ve stem cells determine how the cancer advances.[18] Densitometry results of LGR5 expression by western blotting in the different cell lines showed that high LGR5 expression levels were apparent in BHK, AGS, VERO and NIH3T3 cell lines compared with the other cell lines.[19]
References
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- ↑ McDonald T, Wang R, Bailey W, Xie G, Chen F, Caskey CT, Liu Q (Jul 1998). "Identification and cloning of an orphan G protein-coupled receptor of the glycoprotein hormone receptor subfamily". Biochem Biophys Res Commun. 247 (2): 266–70. doi:10.1006/bbrc.1998.8774. PMID 9642114.
- 1 2 McClanahan T, Koseoglu S, Smith K, Grein J, Gustafson E, Black S, Kirschmeier P, Samatar AA (April 2006). "Identification of overexpression of orphan G protein-coupled receptor GPR49 in human colon and ovarian primary tumors". Cancer Biol. Ther. 5 (4): 419–26. doi:10.4161/cbt.5.4.2521. PMID 16575208.
- 1 2 3 4 Hsu SY, Liang SG, Hsueh AJ (December 1998). "Characterization of two LGR genes homologous to gonadotropin and thyrotropin receptors with extracellular leucine-rich repeats and a G protein-coupled, seven-transmembrane region". Mol. Endocrinol. 12 (12): 1830–45. doi:10.1210/me.12.12.1830. PMID 9849958.
- 1 2 "LGR5 leucine-rich repeat-containing G protein-coupled receptor 5". Entrez Gene.
- ↑ "LGR5 leucine-rich repeat containing G protein-coupled receptor 5". IUPHAR Database.
- ↑ Carmon KS, Lin Q, Gong X, Thomas A, Liu Q (June 2012). "LGR5 interacts and cointernalizes with Wnt receptors to modulate Wnt/β-catenin signaling". Mol. Cell. Biol. 32 (11): 2054–64. doi:10.1128/MCB.00272-12. PMC 3372227
 . PMID 22473993.
. PMID 22473993. - ↑ Morita H, Mazerbourg S, Bouley DM, Luo CW, Kawamura K, Kuwabara Y, Baribault H, Tian H, Hsueh AJ (November 2004). "Neonatal lethality of LGR5 null mice is associated with ankyloglossia and gastrointestinal distension". Mol. Cell. Biol. 24 (22): 9736–43. doi:10.1128/MCB.24.22.9736-9743.2004. PMC 525477. PMID 15509778.
- 1 2 3 4 5 Barker N, Clevers H (May 2010). "Leucine-rich repeat-containing G-protein-coupled receptors as markers of adult stem cells". Gastroenterology. 138 (5): 1681–96. doi:10.1053/j.gastro.2010.03.002. PMID 20417836.
- ↑ Ruffner H, Sprunger J, Charlat O, Leighton-Davies J, Grosshans B, Salathe A, Zietzling S, Beck V, Therier M, Isken A, Xie Y, Zhang Y, Hao H, Shi X, Liu D, Song Q, Clay I, Hintzen G, Tchorz J, Bouchez LC, Michaud G, Finan P, Myer VE, Bouwmeester T, Porter J, Hild M, Bassilana F, Parker CN, Cong F (2012). "R-Spondin potentiates Wnt/β-catenin signaling through orphan receptors LGR4 and LGR5". PLoS ONE. 7 (7): e40976. doi:10.1371/journal.pone.0040976. PMC 3397969. PMID 22815884.
- ↑ Barker N, Rookmaaker MB, Kujala P, Ng A, Leushacke M, Snippert H, van de Wetering M, Tan S, Van Es JH, Huch M, Poulsom R, Verhaar MC, Peters PJ, Clevers H (September 2012). "Lgr5+ve stem/progenitor cells contribute to nephron formation during kidney development". Cell Rep. 2 (3): 540–52. doi:10.1016/j.celrep.2012.08.018. PMID 22999937.
- ↑ Barker N, Huch M, Kujala P, van de Wetering M, Snippert HJ, van Es JH, Sato T, Stange DE, Begthel H, van den Born M, Danenberg E, van den Brink S, Korving J, Abo A, Peters PJ, Wright N, Poulsom R, Clevers H (January 2010). "Lgr5+ve stem cells drive self-renewal in the stomach and build long-lived gastric units in vitro". Cell Stem Cell. 6 (1): 25–36. doi:10.1016/j.stem.2009.11.013. PMID 20085740.
- ↑ Haegebarth A, Clevers H (March 2009). "Wnt signaling, lgr5, and stem cells in the intestine and skin". Am. J. Pathol. 174 (3): 715–21. doi:10.2353/ajpath.2009.080758. PMC 2665733. PMID 19197002.
- ↑ Kleist B, Xu L, Li G, Kersten C (April 2011). "Expression of the adult intestinal stem cell marker Lgr5 in the metastatic cascade of colorectal cancer". Int J Clin Exp Pathol. 4 (4): 327–35. PMC 3093057. PMID 21577318.
- ↑ Schepers AG, Snippert HJ, Stange DE, van den Born M, van Es JH, van de Wetering M, Clevers H (August 2012). "Lineage tracing reveals Lgr5+ stem cell activity in mouse intestinal adenomas". Science. 337 (6095): 730–5. doi:10.1126/science.1224676. PMID 22855427.
- ↑ Walker F, Zhang HH, Odorizzi A, Burgess AW (2011). "LGR5 is a negative regulator of tumourigenicity, antagonizes Wnt signalling and regulates cell adhesion in colorectal cancer cell lines". PLoS ONE. 6 (7): e22733. doi:10.1371/journal.pone.0022733. PMC 3145754. PMID 21829496.
- ↑ Simon E, Petke D, Böger C, Behrens HM, Warneke V, Ebert M, Röcken C (2012). "The spatial distribution of LGR5+ cells correlates with gastric cancer progression". PLoS ONE. 7 (4): e35486. doi:10.1371/journal.pone.0035486. PMC 3329462. PMID 22530031.
- ↑ Alizadeh-Navaei R, Rafiei A, Abedian-Kenari S, Asgarian-Omran H, Valadan R, Hedayatizadeh-Omran A (July 2016). "Comparison of leucine-rich repeat-containing G protein-coupled receptor 5 expression in different cancer and normal cell lines". Biomedical Reports. 5 (1): 130–132. doi:10.3892/br.2016.684. PMC 4907073. PMID 27347416.
Further reading
- Hsu SY, Kudo M, Chen T, Nakabayashi K, Bhalla A, van der Spek PJ, van Duin M, Hsueh AJ (August 2000). "The three subfamilies of leucine-rich repeat-containing G protein-coupled receptors (LGR): identification of LGR6 and LGR7 and the signaling mechanism for LGR7". Mol. Endocrinol. 14 (8): 1257–71. doi:10.1210/me.14.8.1257. PMID 10935549.
- Yamamoto Y, Sakamoto M, Fujii G, Tsuiji H, Kenetaka K, Asaka M, Hirohashi S (March 2003). "Overexpression of orphan G-protein-coupled receptor, Gpr49, in human hepatocellular carcinomas with beta-catenin mutations". Hepatology. 37 (3): 528–33. doi:10.1053/jhep.2003.50029. PMID 12601349.
This article incorporates text from the United States National Library of Medicine, which is in the public domain.