Cerebral organoid
A cerebral organoid describes artificially grown, in vitro, miniature organs resembling the brain. Cerebral organoids are created by culturing human pluripotent stem cells in a three-dimensional rotational bioreactor and develop over a course of months.[1] The human brain is an extremely complex system of heterogeneous tissues and consists of an extremely diverse array of neurons. This complexity has made studying the brain and understanding how it works a difficult task in neuroscience, especially when it comes to neurodegenerative diseases. The purpose of creating an in vitro neurological model is to study these diseases in a more simple and variable space; free of in vivo limitations, especially when working with humans. The varying physiology between human and other mammalian models limits the scope of study in neurological disorders. Cerebral organoids are synthesized tissues that contain several types of nerve cells and have anatomical features that resemble mammalian brains. Cerebral organoids are most similar to layers of neurons called the cortex and choroid plexus. In some cases, structures similar to the retina, meninges and hippocampus can form.[1][2] Stem cells have the potential to grow into many different types of tissues and their fate is dependent on many factors. Below is an image showing some of the chemical factors that can lead stem cells to differentiate into various neural tissues. Similar techniques are used on stem cells used to grow cerebral organoids.[3]
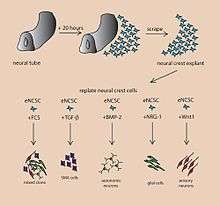
Model Development
Using human pluripotent stem cells to create in vitro cerebral organoids allows researchers to summarize current developmental mechanisms for human neural tissue as well as study the roots of human neurological diseases. Cerebral organoids are an investigative tool, used to understand how disease pathology works. These organoids can be used in experiments that current in vitro methods are too simple for, while also being more human applicable than rodent or other mammalian models. Historically, major breakthroughs in how the brain works have resulted from injury or disorder in human brain function, leading to understanding of how regions of the brain work. An in vitro human brain model would allow for the next wave in understanding of the human brain.[1][3]
Applications
In addition to being used as tools to study disease pathology and treatments, future application of cerebral organoids include direct implantation into a host, like a human being. The organoid can fuse with host tissue in areas of neurodegeneration, being incorporated with the host vasculature and be immunologically silent.[4] A list of potential applications for cerebral organoids is highlighted below.
Potential Applications
- Tissue morphogenesis with respect to cerebral organoids covers how neural organs form in vertebrates. Cerebral organoids can serve as in vitro tools to study the formation, modulate it, and further understand the mechanisms controlling it.[5]
- Migration Assays
- Cerebral organoids can help to study cell migration. Neural glial cells cover a wide variety of neural cells, some of which move around the neurons. The factors that govern their movements can be studied using cerebral organoids.[3]
- Clonal lineage tracing
- Clonal lineage tracing is part of fate mapping, where the lineage of differentiated tissues is traced to the pluripotent progenitors. The local stimuli released and mechanism of differentiation can be studied using cerebral organoids as a model.[5]
- Transplantation
- Cerebral organoids can be used to grow specific brain regions and transplant them into regions of neurodegeneration as a therapeutic treatment.
- Cell fate potential
- Cross species developmental timing
- Cerebral organoids provide a unique insight into the timing of development of neural tissues and can be used as a tool to study the differences across species.[5]
- Cerebral organoids can be used as simple models of complex brain tissues to study the effects of drugs and to screen them for initial safety and efficacy.
- Cell replacement therapy
- Cerebral organoids can be used as a simple model to show how cell replacement therapy would work on brain tissues.[5]
- Cell-type specific genome assays
Disease
Organoids can be used to study the crucial early stages of brain development, test drugs and, because they can be made from living cells, study individual patients.,[2] In one case, a cerebral organoid grown from a patient with microcephaly demonstrated related symptoms and revealed that apparently the cause is overly rapid development, followed by slower brain growth. Microencephaly is a developmental condition in which the brain remains undersized, producing an undersized head and debilitation. Microcephaly is not suitable for mouse models, which do not replicate the condition.[2] The primary form of the disease is thought to be caused by a homozygous mutation in the microcephalin gene. The disease is difficult to reproduce in mouse models because mice lack the developmental stages for an enlarged cerebral cortex that humans have. Naturally, a disease which affects this development would be impossible to show in a model which does not have it to begin with.[6] To use cerebral organoids to model a human's microcephaly, one group of researchers has taken patient skin fibroblasts and reprogrammed them using four well known reprogramming factors. These include OCT4, SOX2, MYC and KLF4. The reprogrammed sample was able to be cloned into induced pluripotent stem cells. The cells were cultured into a cerebral organoid following a process described in the cerebral organoid creation section below. The organoid that resulted had decreased numbers of neural progenitor cells and smaller tissues. Additionally, the patient derived tissues displayed fewer and less frequent neuroepithelial tissues made of progenitors, decreased radial glial stem cells, and increased neurons. These results suggest that the underlying mechanism of microcephaly is caused by cells prematurely differentiating into neurons leaving a deficit of radial glial cells.[1]
Construction
To make an organoid, an embryoid (tissue that has some embryonic features) grown from natural stem cells is used. Embryos have three layers: endoderm, mesoderm and ectoderm. Each turns into various body parts. The nervous system grows from the ectoderm (which also contributes dental enamel and the epidermis).[3] Ectodermal cells were placed into gel droplets and floated in a nutrient broth in a rotating bioreactor, which supported cell growth without forming by the container. After ten days the organoid developed neurons. After 30 days it displayed regions similar to parts of brains. Lacking a blood supply, cerebral organoids reach about 4 mm across and can last a year or more.[2] The general procedure can be broken down into 5 steps. First human pluripotent stem cells are cultured. They are then allowed to cultivate into an embryoid body. Next the cell culture is induced to form a neuroectoderm. The neuroectoderm is them grown in a matrigel droplet. The matrigel provides nutrients and the neuroectoderm starts to proliferate and grow. However, the lack of vasculature limits the size the organoid can grow. This has been the major limitation in organoid development, however new methods using a spinning bioreactor have allowed an increase in the availability of nutrients to cells inside the organoid. This last step has been the key breakthrough in organoid development.[7] Spinning bioreactors have been used increasingly in cell culture and tissue growth applications. The reactor is able to deliver faster cell doubling times, increased cell expansion and increased extra-cellular matrix components when compared to statically cultured cells.[8]
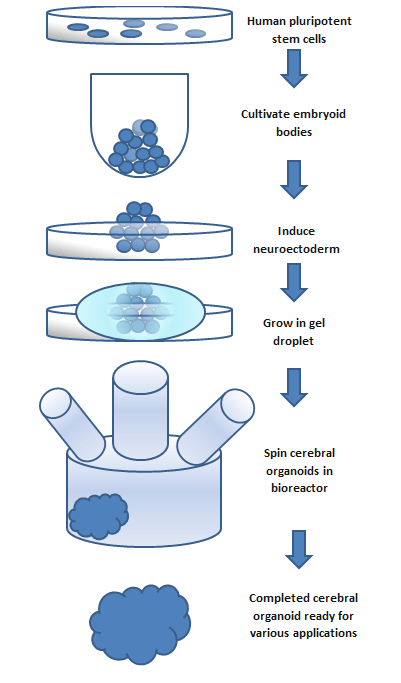
Figure: This flow chart outlines the basic steps to create a cerebral organoid. The process takes a span of months and the size of the organoid is limited to the availability of nutrients.
Testing
Differentiation
It has been shown that cerebral organoids grown using the spinning bioreactor 3D culture method differentiate into various neural tissue types, such as the optic cup, hippocampus, ventral parts of the teleencephelon and dorsal cortex.[9] The neural stem/progenitor cells are unique because they are able to self-renew and are multipotent. This means they can generate neurons and glial cells which are the two main components of neural systems. The fate of these cells is controlled by several factors that affect the differentiation process. The spatial location and temporal attributes of neural progenitor cells can influence if the cells form neurons or glial cells. Further differentiation is then controlled by extracellular conditions and cell signaling.[10] The exact conditions and stimuli necessary to differentiate neural progenitor cells into specific neural tissues such as hippocampal tissue, optic nerve, cerebral cortex, etc. are unknown. It is believed that cerebral organoids can be used to study the developmental mechanisms of these processes.[7]
Gene Expression
To test if the neural progenitor cells and stem cells are differentiating into specific neural tissues, several gene markers can be tested. Two markers that are present during pluripotent stages are OCT4 and NANOG. These two markers are diminished during the course of development for the organoid. Neural identity markers that note successful neural induction, SOX1 and PAX6, are upregulated during organoid development. These changes in expression support the case for self-guided differentiation of cerebral organoids.[1] Markers for forebrain and hindbrain can also be tested. Forebrain markers FOXG1 and SIX3 are highly expressed throughout organoid development. However, hindbrain markers EGR2 and ISL1 show early presence but decrease in the later stages. This imbalance towards forebrain development is similar to developmental expansion of forebrain tissue in human brain development.[1] To test if organoids develop even further into regional specification, gene markers for cerebral cortex and occipital lobe have been tested. Many regions that have forebrain marker FOXG1, labeling them as regions with cerebral cortical morphology, were also positive for marker EMX1 which indicates dorsal cortical identity. These specific regions can be even further specified by markers AUTS2, TSHZ2, and LMO4 with the first representing cerebral cortex and the two after representing the occipital lobe.[1] Genetic markers for the hippocampus, ventral forebrain, and choroid plexus are also present in cerebral organoids, however the overall structures of these regions have not yet been formed.
Localization
Functional
Cerebral organoids also possess functional cerebral cortical neurons. These neurons must form on the radially organized cortical plate. The marker TBR1 is present in the preplate, the precursor to the cortical plate, and is present, along with MAP2, a neuronal marker, in 30-day-old cerebral organoids. These markers are indicative of a basal neural layer similar to a preplate. These cells are also apically adjacent to a neutral zone and are reelin+ positive, which indicates the presence of Cajal-Retzius cells. The Cajal-Retzius cells are important to the generation of cortical plate architecture.[7] The cortical plate is usually generated inside-out such that later born neurons migrate to the top superficial layers. This organization is also present in cerebral organoids based on genetic marker testing. Neurons that are early born have marker CTIP2 and are located adjacent to the TBR1 exhibiting preplate cells. Late born neurons with markers SATB2 and BRN2 are located in a superficial layer, further away from the preplate than the early born neurons suggesting cortical plate layer formation. Additionally, after 75 days of formation, cerebral organoids show a rudimentary marginal zone, a cell-poor region. The formation of layered cortical plate is very basic in cerebral organoids and suggests the organoid lacks the cues and factors to induce formation of layer II-VI organization.[1] The cerebral organoid neurons can, however form axons as shown by GFP staining. GFP labeled axons have been shown to have complex branching and growth cone formation. Additionally calcium dye imaging has shown cerebral organoids to have Ca2+ oscillations and spontaneous calcium surges in individual cells. The calcium signaling can be enhanced through glutamate and inhibited through tetrodotoxin.[1]
Physical
It is not fully understood how individual localized tissues formed by stem cells are able to coordinate with surrounding tissues to develop into a whole organ.[11] It has been shown however that most tissue differentiation requires interactions with surrounding tissues and depends on diffusible induction factors to either inhibit or encourage various differentiation and physical localization.[11] Cerebral organoid differentiation is somewhat localized. The previously mentioned markers for forebrain and hindbrain are physically localized, appearing in clusters. This suggests that local stimuli are released once one or more cells differentiates into a specific type as opposed to a random pathway throughout the tissue. The markers for subspecification of cortical lobes, prefrontal cortex and occipital lobe, are also physically localized. However, the hippocampus and ventral forebrain cells are not physically localized and are randomly located through the cerebral organoid.[1] Cerebral organoids lack blood vessels and are limited in size by nutrient uptake in the innermost cells. Spinning bioreactors and advanced 3D scaffolding techniques are able to increase organoid size, though the integration of in vitro nutrient delivery systems is likely to spark the next major leap in cerebral organoid development.[5]
References
- 1 2 3 4 5 6 7 8 9 10 Lancaster, M. A.; Renner, M.; Martin, C. A.; Wenzel, D.; Bicknell, L. S.; Hurles, M. E.; Homfray, T.; Penninger, J. M.; Jackson, A. P.; Knoblich, J. A. (2013). "Cerebral organoids model human brain development and microcephaly". Nature. doi:10.1038/nature12517.
- 1 2 3 4 "Growing model brains: An embryonic idea". The Economist. 2013-08-31. Retrieved 2013-09-07.
- 1 2 3 4 al.], edited by Dale Purves ... [et (2007). Neuroscience. (4th ed.). New York: W. H. Freeman. ISBN 0878936971.
- ↑ Lanza, edited by Anthony Atala, Robert P. (2002). Methods of tissue engineering ([Nachdr.]. ed.). San Diego, CA: Academic Press. p. 381. ISBN 9780124366367.
- 1 2 3 4 5 6 Chambers, Stuart M.; Tchieu, Jason; Studer, Lorenz (October 2013). "Build-a-Brain". Cell Stem Cell. 13 (4): 377–378. doi:10.1016/j.stem.2013.09.010. PMID 24094317.
- ↑ Opitz, JM; Holt, MC (1990). "Microcephaly: general considerations and aids to nosology.". Journal of craniofacial genetics and developmental biology. 10 (2): 175–204. PMID 2211965.
- 1 2 3 Vogel, Gretchen (30 August 2013). "Lab Dishes Up Mini-Brains". Science. doi:10.1126/science.341.6149.946. Retrieved 1 November 2013.
- ↑ Lueders, Cora (14 June 2013). "Large Scale Expansion of Human Umbilical Cord Cells in a Rotating Bed System Bioreactor for Cardiovascular Tissue Engineering Applications". The Open Biomedical Engineering Journal. 7 (1): 50–61. doi:10.2174/1874120701307010050.
- ↑ Bershteyn, Marina; Kriegstein, Arnold R. (September 2013). "Cerebral Organoids in a Dish: Progress and Prospects". Cell. 155 (1): 19–20. doi:10.1016/j.cell.2013.09.010.
- ↑ Sakayori, N; Kikkawa, T; Osumi, N (Oct 2012). "Reduced proliferation and excess astrogenesis of Pax6 heterozygous neural stem/progenitor cells.". Neuroscience research. 74 (2): 116–21. doi:10.1016/j.neures.2012.08.004. PMID 22944581.
- 1 2 Eiraku, Mototsugu; Takata, Nozomu; Ishibashi, Hiroki; Kawada, Masako; Sakakura, Eriko; Okuda, Satoru; Sekiguchi, Kiyotoshi; Adachi, Taiji; Sasai, Yoshiki (7 April 2011). "Self-organizing optic-cup morphogenesis in three-dimensional culture". Nature. 472 (7341): 51–56. doi:10.1038/nature09941. PMID 21475194.