Axoplasmic transport
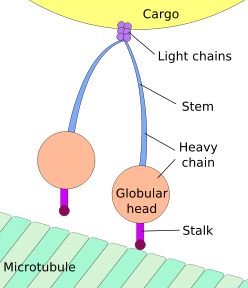
Axoplasmic transport, also called axonal transport, is a cellular process responsible for movement of mitochondria, lipids, synaptic vesicles, proteins, and other cell parts (i.e. organelles) to and from a neuron's cell body, through the cytoplasm of its axon (the axoplasm). Axons, which can be 1,000 or 10,000 times the length of the cell body, were originally thought not to contain any ribosomes as the means of producing proteins, and so were thought to rely on axoplasmic transport for all their protein needs.[1][2] However, more recently, mRNA translation has been demonstrated in axons.[3][4] Axonal transport is also responsible for moving molecules destined for degradation from the axon back to the cell body, where they are broken down by lysosomes.[5]
Movement toward the cell body is called retrograde transport and movement toward the synapse is called anterograde transport.[1][6][7]
Mechanism
The vast majority of axonal proteins are synthesized in the neuronal cell body and transported along axons. Axonal transport occurs throughout the life of a neuron and is essential to its growth and survival. Microtubules (made of tubulin) run along the length of the axon and provide the main cytoskeletal "tracks" for transportation. Kinesin and dynein are motor proteins that move cargoes in the anterograde (forwards from the soma to the axon tip) and retrograde (backwards to the soma (cell body)) directions, respectively. Motor proteins bind and transport several different cargoes including mitochondria, cytoskeletal polymers, autophagosomes, and synaptic vesicles containing neurotransmitters.[1]
Axonal transport can be fast or slow, and anterograde or retrograde.
Fast and slow transport
Vesicular cargoes move relatively fast (50–400 mm/day) whereas transport of soluble (cytosolic) and cytoskeletal proteins takes much longer (moving at less than 8 mm/day). The basic mechanism of fast axonal transport has been understood for decades but the mechanism of slow axonal transport is only recently becoming clear, as a result of advanced imaging techniques.[8] Fluorescent labeling techniques (e.g. fluorescence microscopy) have enabled direct visualization of transport in living neurons. (See also: Anterograde tracing.)
Recent studies have revealed that the movement of cytoskeletal "slow" cargoes is actually rapid but unlike fast cargoes, they pause frequently, making the overall transit rate much slower. The mechanism is known as the "Stop and Go" model of slow axonal transport, and has been extensively validated for the transport of the cytoskeletal protein neurofilament.[9] The movement of soluble (cytosolic) cargoes is more complex, but appears to have a similar basis where soluble proteins organize into multi-protein complexes that are then conveyed by transient interactions with more rapidly moving cargoes moving in fast axonal transport.[10][11][12] An analogy is the difference in transport rates between local and express subway trains. Though both types of train travel at similar velocities between stations, the local train takes much longer to reach the end of the line because it stops at every station whereas the express makes only a few stops on the way.
Anterograde transport
Anterograde (also called "orthograde") transport is movement of molecules/organelles outward, from the cell body (also called soma) to the synapse or cell membrane.
The anterograde movement of individual cargoes (in transport vesicles) of both fast and slow components along the microtubule[7] is mediated by kinesins. Several kinesins have been implicated in slow transport,[8] though the mechanism for generating the "pauses" in the transit of slow component cargoes is still unknown.
There are two classes of slow anterograde transport: slow component a (SCa) that carries mainly microtubules and neurofilaments at 0.1-1 millimeters per day, and slow component b (SCb) that carries over 200 diverse proteins and actin at a rate of up to 6 millimeters per day.[8] The slow component b, which also carries actin, are transported at a rate of 2-3 millimeters per day in retinal cell axons.
During reactivation from latency, the herpes simplex virus (HSV) enters its lytic cycle, and uses anterograde transport mechanisms to migrate from dorsal root ganglia neurons to the skin or mucosa that it subsequently affects.[13]
Retrograde transport
Retrograde transport is movement of molecules/organelles inward, away from the synapse or plasma membrane toward the cell body or soma. Retrograde transport is mediated by dynein, and is used for example to send chemical messages and endocytosis products headed to endolysosomes from the axon back to the cell.[5] Fast retrograde transport can cover 10-20 centimeters per day.[5]
Fast retrograde transport returns used synaptic vesicles and other materials to the soma and informs the soma of conditions at the axon terminals. Some pathogens exploit this process to invade the nervous system. They enter the distal tips on an axon and travel to the soma by retrograde transport. Examples include tetanus toxin and the herpes simplex, rabies, and polio viruses. In such infections, the delay between infection and the onset of symptoms corresponds to the time needed for the pathogens to reach the somata.[14]
Consequences of interruption
Several rare neurodegenerative diseases are linked to genetic mutations in the motor proteins, kinesin and dynein, and in those cases it is likely that axonal transport is a key player in mediating pathology. Dysfunctional axonal transport is also linked to sporadic (common) forms of neurodegenerative diseases such as Alzheimer's and Parkinson's.[8] This is mainly due to numerous observations that large axonal accumulations are invariably seen in affected neurons, and that genes known to play a role in the familial forms of these diseases also have purported roles in normal axonal transport. However, there is little direct evidence for involvement of axonal transport in the latter diseases, and other mechanisms (such as direct synaptotoxicity) may be more relevant.
Since the axon depends on axoplasmic transport for vital proteins and materials, injury such as diffuse axonal injury that interrupts the transport will cause the distal axon to degenerate in a process called Wallerian degeneration. Cancer drugs that interfere with cancerous growth by altering microtubules (which are necessary for cell division) damage nerves because the microtubules are necessary for axonal transport.[1]
Infection
The rabies virus reaches the central nervous system by retrograde axoplasmic flow.[15]
References
- 1 2 3 4 Cowie R.J. and Stanton G.B. "Axoplasmic Transport and Neuronal Responses to Injury." Howard University College of Medicine. Retrieved on January 25, 2007.
- ↑ Sabry, J.; O'Connor, T. P.; Kirschner, M. W. (1995). "Axonal Transport of Tubulin in Ti1 Pioneer Neurons in Situ". Neuron. 14 (6): 1247–1256. doi:10.1016/0896-6273(95)90271-6. PMID 7541635.
- ↑ Giustetto, M; Hegde, AN; Si, K; et al. (2003). "Axonal transport of eukaryotic translation elongation factor 1alpha mRNA couples transcription in the nucleus to long-term facilitation at the synapse". Proc. Natl. Acad. Sci. USA. 100 (23): 13680–5. doi:10.1073/pnas.1835674100. PMC 263873
 . PMID 14578450.
. PMID 14578450. - ↑ Si K, Giustetto Etkin, A; et al. (Dec 2003). "A neuronal isoform of CPEB regulates local protein synthesis and stabilizes synapse-specific long-term facilitation in aplysia". Cell. 115 (7): 893–904. doi:10.1016/s0092-8674(03)01021-3. PMID 14697206.
- 1 2 3 Oztas, E (2003). "Neuronal Tracing." (PDF). Neuroanatomy. 2: 2–5.
- ↑ Karp G. 2005. Cell and Molecular Biology: Concepts and Experiments, Fourth ed, p. 344. John Wiley and Sons, Hoboken, NJ. ISBN 0-471-46580-1
- 1 2 Bear et al., 2006. "Neuroscience: Exploring the Brain," 3/e, p. 41
- 1 2 3 4 Roy, S; et al. (2005). "Axonal transport defects: a common theme in neurodegenerative". Acta Neuropathol. 109 (1): 5–13. doi:10.1007/s00401-004-0952-x. PMID 15645263.
- ↑ Brown 2003. "Axonal transport of membranous and nonmembranous cargoes: a unified perspective", J Cell Biol. 2003 Mar 17;160(6):817-21
- ↑ Scott et al., 2011. Mechanistic logic underlying the axonal transport of cytosolic proteins". Neuron. 2011 May 12;70(3):441-54.
- ↑ Roy S et al., 2007. "Rapid and intermittent cotransport of slow component-b proteins". J Neurosci. 2007 Mar 21;27(12):3131-8
- ↑ Kuznetsov, Andrey V. (2011). "Analytical solution of equations describing slow axonal transport based on the stop-and-go hypothesis". Central European Journal of Physics. 9 (3): 662–673. doi:10.2478/s11534-010-0066-0.
- ↑ Holland, David J.; Miranda-Saksena, Monica; et al. (1999). "Anterograde Transport of Herpes Simplex Virus Proteins in Axons of Peripheral Human Fetal Neurons: an Immunoelectron Microscopy Study". Journal of Virology. 73 (10): 8503–8511. PMC 112870. PMID 10482603.
- ↑ Saladin, Kenneth. Anatomy and Physiology: The Unity of Form and Function. Sixth. New York : McGraw-Hill, 2010. 445. Print.
- ↑ Mitrabhakdi, E; Shuangshoti, S; Wannakrairot, P; Lewis, RA; Susuki, K; et al. (2005). "Differences in neuropathogenetic mechanisms in human furious and paralytic rabies". J Neurol Sci. 238: 3–10. doi:10.1016/j.jns.2005.05.004. PMID 16226769.